|
|
НОВЫЕ ВИДЫ ОБРАЗУЮТСЯ ОЧЕНЬ БЫСТРО, ЕСЛИ МЕНЯЕТСЯ СИСТЕМА РАСПОЗНАВАНИЯ ПОЛОВЫХ ПАРТНЕРОВ
Конкуренция и другие межвидовые взаимодействия могут стимулировать быстрое видообразование, когда к ним присоединяется естественный отбор, направленный на совершенствование механизмов, обеспечивающих успешную встречу брачных партнеров в пределах одного из видов. К такому выводу пришли австралийские ученые на основании изучения разных природных популяций одного вида тропических древесных лягушек.
Рис 1. Обитающая во влажных тропических лесах Австралии зеленоглазая древесная лягушка Litoria genimaculata служила объектом исследования для Хоскина и Хигги. Снимки © James Reardon с сайта jamesreardon.wordpress.com
Еще Чарльз Дарвин обратил внимание на то, что в некоторых, богатых видами, систематических группах (родах, семействах), наиболее близкородственные виды различаются прежде всего признаками, связанными с выбором брачного партнера и осуществлением спаривания. Это могут быть как особенности строения половых органов, так и специфические сигналы для привлечения партнера — к примеру, кваканье самца лягушки или пахучие вещества (аттрактанты), выделяемые самками (а иногда самцами) некоторых насекомых. Если в какой-то части популяции идет отбор на изменение таких признаков, то она может оказаться репродуктивно изолированной (иногда говорят — «сексуально изолированной») от материнской популяции. Особенно быстро это происходит тогда, когда гибриды между особями из отделившейся группы и материнской популяции отличаются пониженной жизнеспособностью. Такое нарастание степени сексуальной изоляции и, соответственно, ускорение образования нового вида получило в англоязычной литературе специальное наименование — «reinforcement» (что буквально означает «усиление»). В настоящее время специалисты в области эволюционной биологии интенсивно изучают «reinforcement» и часто связывают его с таким давно описанным явлением, как «смещение признаков» (character displacement) — изменение признака не у всех популяций вида, а только у тех, которые соприкасаются с экологически близкими конкурирующими видами. Так, у двух видов галапагосских вьюрков, обитающих на разных островах (в условиях аллопатрии), клювы примерно одинакового размера, но когда те же самые виды обитают на одном острове (в условиях симпатрии), то средние размеры их клювов существенно различаются, что заставляет предположить и наличие различий в потребляемой ими пище. Если смещающийся при наличии конкурента признак ответственен за обеспечение встречи с половым партнером, то это может привести к очень быстрому видообразованию, поскольку прекращается обмен генами между отпочковывающейся и материнской популяциями.
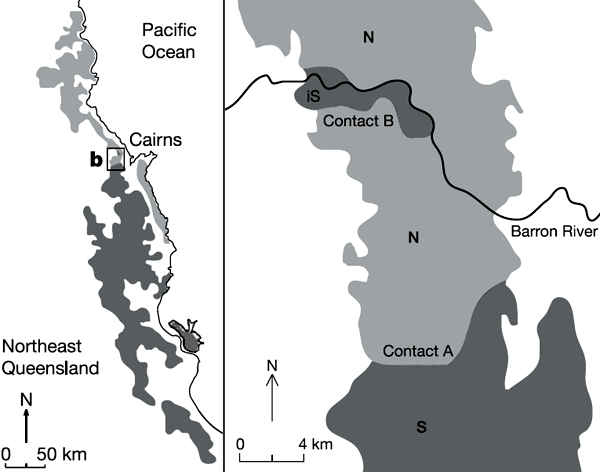
В журнале «Ecology Letters» недавно появилась обзорная статья двух австралийских исследователей, Конрада Хоскина (Conrad Hoskin) и Меган Хигги (Megan Higgie), которые подробно разбирают каким образом от популяции одного вида посредством изменения признаков, отвечающих за брачное поведение, может отделиться новый вид. Объект их исследования — зеленоглазая древесная лягушка (Green-eyed tree frog), Litoria genimaculata (рис. 1), вид, довольно широко распространенный во влажных тропических лесах Австралии и Новой Гвинеи. Там, где непосредственно проводили работы, в районе реки Баррон (Barron, северо-восточная часть штата Квинсленд, Австралия), существуют две (это доказано молекулярно-генетическими методами) аллопатрические — то есть занимающие разные территории — популяции зеленоглазых лягушек, условно обозначенные как «северная» и «южная» (рис. 2). Сформировались эти популяции, видимо, когда-то в плейстоцене, в период более холодного и засушливого климата, когда тропические леса оставались только в виде отдельных островов среди открытого ландшафта.
Рис 2. Район исследований (северо-восточная часть Квинсленда, Австралия). Справа показаны районы обитания «южной» (S — выделено темно-серым) и «северной» (N — выделено светло-серым) популяций зеленоглазой древесной лягушки. Обратите внимание на анклав «южной» популяции в северной части района обитания «северной» популяции. Именно здесь и шло интенсивное смещение признаков, обеспечивающих встречу брачных партеров. Рис. из статьи: Hoskin et al. // Nature. 2005. V. 437. P. 1353–1356
Позднее, примерно 6500 лет назад, когда климат стал более теплым и влажным, а тропические леса снова образовали сплошной покров, зоны распространения этих популяций лягушек сомкнулись. Гибриды между ними в пограничной области возможны, хотя они характеризуются пониженной жизнеспособностью. Своеобразная ситуация сложилась, однако, на севере обследованной территории, где в окружении «северной» популяции оказался небольшой «анклав», представленный особями «южной» популяции (на рис. 2 он помечен как iS). Именно в этом «анклаве» и было обнаружено необычайно быстрое смещение признаков, отвечающих за встречу полов (характеристики призывного крика самцов и ответа самки). В результате этого смещения особи данной группировки утратили способность к нормальному скрещиванию с особями материнской (основной «южной» популяции). Как это могло происходить, схематически показано на приведенных ниже рисунках 3 и 4.
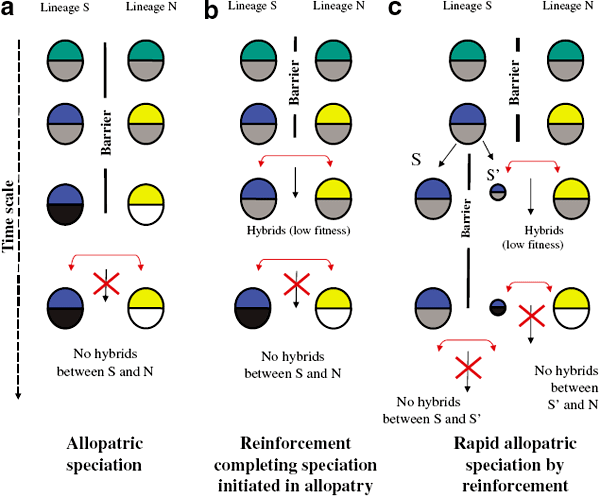
Рис. 3. Схема видообразования на примере расхождения (и утраты способности образовывать гибриды) двух генетически различающихся линий в одной популяции — «южной» (S) и «северной» (N). Время движется сверху вниз. Рассмотрены три варианта: a — классическое аллопатрическое видообразование; b — начавшееся аллопатрическое видообразование завершается «reinforcement» (наступлением стерильности или гибели гибридов); c — быстрое аллопатрическое видообразование, усиленное с самого начала «reinforcement» и приведшее к появлению еще одной самостоятельной генетической линии (S’), фактически — к возникновению третьего вида. Подробности в тексте. Рис. из статьи: Smadja C. & Butlin R. // Heredity. 2006. V. 96. P. 422–423
Рис. 3 иллюстрирует три возможных варианта эволюционных изменений в отдельных генетических линиях популяции: в «южной» (S) и «северной» (N). Каждый кружок соответствует определенной линии в определенный момент времени. Верхняя половинка кружка — это постзиготическая (наступающая после оплодотворения) изоляция. Если для сосуществующих популяций эти половинки одного цвета, то гибриды жизнеспособны, если разного — нежизнеспособны. Нижняя половинка кружка соответствует презиготической (то есть существующей еще до оплодотворения) изоляции: спаривание особей разных линий возможно, если они одного цвета, невозможно — если разного. Шкала времени (и, соответственно, эволюции) направлена сверху вниз. Барьеры, мешающие встрече особей разных генетических линий, показаны черными вертикальными линиями и обозначены словом «Barrier». Красные горизонтальные стрелки показывают возможность контакта особей разных линий. Черная тонкая стрелка вниз — скрещивание особей разных линий. Красный крест показывает невозможность гибридизации. Рассмотрены три случая: a — классическое аллопатрическое видообразование (постепенное накопление различий в изолированных друг от друга популяциях и утрата способности к скрещиванию); b — начавшееся аллопатрическое видообразование завершается «reinforcement» (хотя особи еще способны спариваться, их гибриды стерильны или отличаются пониженной жизнеспособностью); c — начавшееся аллопатрическое видообразование меняет свой ход из-за новой системы барьеров: популяция «южной» линии оказывается подразделенной новым барьером, при этом часть ее попадает по одну сторону барьера вместе с «северной» линией; именно в этой части южной популяции (показана небольшим кружком с индексом S’) идет отбор, направленный на то, чтобы как можно быстрее изолироваться от «северной» популяции (это возможно за счет поведенческих механизмов, обеспечивающих встречу брачных партнеров); при этом отбор заходит так далеко, что особи новой линии (S’) теряют способность скрещиваться с особями материнской южной линии (S), фактически приводя к образованию нового вида.
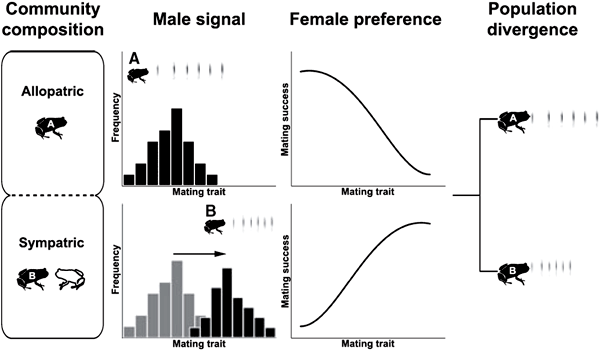
Рис. 4. Схема процесса видообразования, идущего посредством «смещения репродуктивных признаков». Рассматриваемый широко распространенный вид лягушек (выделен черным цветом) в одной части ареала (популяция A; верхние рисунки) не встречается с близко родственным видом (выделен белым цветом), а в другой — встречается (популяция B; нижние рисунки). Соответственно, говорят, что популяция A черного вида аллопатрична по отношению к белому виду, а популяция B — симпатрична. Распределение выраженности признака — звукового призывного звука самца (male signal) — показано гистограммой. При наличии по соседству другого вида, издающего похожие звуки, всё распределение сдвигается вправо, так, чтобы отличаться от конкурента. Также меняется при наличии рядом другого вида и предпочтение самок в выборе самца (female preference). Популяции A и B, хотя и относятся к одному виду, начинают таким образом резко расходиться по признакам, обеспечивающим образование брачных пар. Не исключено, что они потеряют, если уже не потеряли способность к скрещиванию, то есть стали самостоятельными видами. Рис. из обсуждаемой статьи Hoskin & Higgie из Ecology Letters
На рис. 4. более подробно проиллюстрирован как раз вариант смещения репродуктивных признаков в популяции лягушек A в случае обитания рядом с конкурирующим видом. Отбор по репродуктивным признакам, направленный на усиление отличий от конкурента, приводит к тому, что особи данной части популяции (B) могут утерять возможность скрещивания с родительской популяцией и, по сути, дать новый вид. Таким образом, межвидовые взаимодействия могут стимулировать видообразование в пределах одного из видов. Хоскин и Хигги в своем обзоре описывают и других типы межвидовых взаимодействий, которые могут приводить к смещению репродуктивных признаков. Это и воздействие на популяцию хищников и паразитов, которые находят своих жертв, ориентируясь на сигналы, подаваемые теми для привлечения брачных партнеров. Это и опыление, когда некоторые орхидеи начинают образовывать пахучие вещества, напоминающие половые аттрактанты насекомых-опылителей, в результате чего те «спариваются» с цветками вместо того, чтобы спариваться с самками (см. рис. 5). Соответственно, отбор у насекомых-опылителей должен идти на выработку отличий аттрактантов от тех симуляторов, которые производят орхидеи. Помимо этого, авторы указывают на возможность смещения репродуктивных признаков в результате гнездового паразитизма у птиц (известного всем на примере кукушки), мимикрии (к примеру, подражанию ядовитым видам), и даже конкуренции за «сигнальное пространство» (проблема «шумных соседей»).
Рис. 5. Серия снимков, показывающая, как самец перепончатокрылого Neozeleboria cryptoides, принимая за самку специальные выросты цветка орхидеи Chiloglottis trapeziformis, пытается спариваться с цветком и при этом переносит комочек пыльцы (поллинарий) с одного цветка на другой. Вверху слева — самец пока еще на стебле орхидеи. Вверху справа — пытается спариваться с цветком (принимает за бескрылую самку своего вида темные выросты лепестка). Внизу слева — самец уже с прикрепившимися к нему ярко-желтыми поллинариями орхидеи. Внизу справа — готов перелететь на следующий цветок. Самка перепончатокрылого при этом остается неоплодотворенной. Отбор у перепончатокрылых направлен на то, чтобы изменить пахучие вещества (феромоны) самки, привлекающие самцов, и помешать обману со стороны орхидей. Снимки профессора Рода Пиколла (Rod Peakall) с сайта www.anu.edu.au
Обнаружить в природе смещение репродуктивных признаков одного вида (которое может привести к образованию нового вида), вызванное взаимодействиями с другими видами, на самом деле очень непросто. И одно из затрудняющих обстоятельств — необычайная быстрота этих изменений. Результаты исследований австралийских ученых — еще одно подтверждение того, что наша невозможность увидеть воочию процесс видообразования связана не с тем, что процесс этот чрезвычайно длительный (как полагали во времена Дарвина), а наоборот — очень быстрый. 26-04-2010 | Просмотров: 11061
Комментарии
Комментировать
Комментировать
|

Ещё по теме
|
|
|