|
|
ПОЧЕМУ РАЗДЕЛЬНОПОЛОСТЬ ПРЕОБЛАДАЕТ В ПРИРОДЕ?Самооплодотворяющиеся животные размножаются при прочих равных вдвое быстрее, чем раздельнополые. Почему же раздельнополость преобладает в природе? Для ответа на этот вопрос были искусственно выведены породы круглых червей Caenorhabditis elegans, одни из которых практикуют только перекрестное оплодотворение, другие — только самооплодотворение. Эксперименты с этими червями подтвердили две гипотезы о преимуществах перекрестного оплодотворения. Одно преимущество состоит в более эффективном очищении генофонда от вредных мутаций, второе — в ускоренном накоплении полезных мутаций, что помогает популяции приспосабливаться к меняющимся условиям.
Двойная цена самцовЗачем нужно половое размножение, зачем нужны самцы? Ответы на эти вопросы совсем не так очевидны, как может показаться.
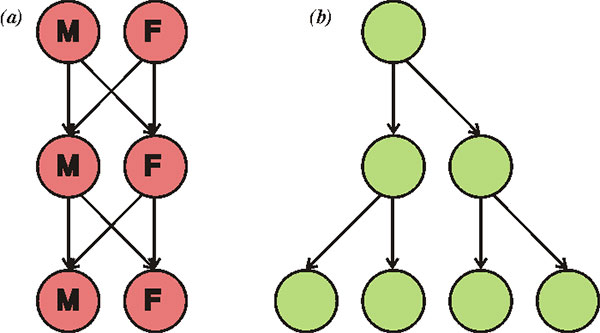
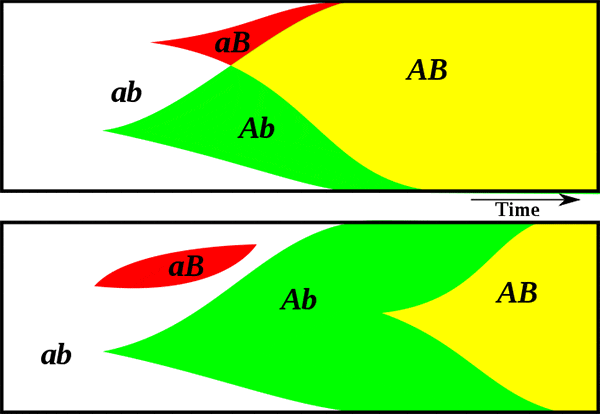
Известный эволюционист Джон Мэйнард Смит (John Maynard Smith) обратил внимание на серьезность данной проблемы в своей книге The Evolution of Sex (1978). Мэйнард Смит подробно рассмотрел парадокс, которому дал название «двойная цена пола» (two-fold cost of sex). Суть его в том, что при прочих равных бесполое размножение (или самооплодотворение) ровно в два раза эффективнее, чем перекрестное оплодотворение с участием самцов (см. рисунок). Иными словами, самцы обходятся популяции непомерно дорого. Отказ от них дает немедленный и очень значительный выигрыш в скорости размножения. Мы знаем, что чисто технически переход от раздельнополости и перекрестного оплодотворения к бесполому размножению или самооплодотворению вполне возможен, тому есть масса примеров как у растений, так и у животных (см., например: Самки гигантского комодского варана размножаются без участия самцов, «Элементы», 26.12.2006). Тем не менее бесполые расы и популяции самооплодотворяющихся гермафродитов почему-то до сих пор так и не вытеснили тех, кто размножается «обычным» образом, с участием самцов. Зачем они все-таки нужны?Из сказанного следует, что перекрестное оплодотворение должно давать некие преимущества, настолько значительные, что они перекрывают даже двойной выигрыш в эффективности размножения, даваемый отказом от самцов. Причем эти преимущества должны проявляться сразу, а не когда-нибудь через миллион лет. Естественному отбору нет дела до отдаленных перспектив. Есть много гипотез о природе этих преимуществ (см.: Evolution of sexual reproduction). Мы рассмотрим две из них. Первая известна под названием «храповик Мюллера» (см.: Muller's ratchet). Храповик — это устройство, в котором ось может крутиться только в одну сторону. Суть идеи в том, что если у бесполого организма возникает вредная мутация, его потомки уже не могут от нее избавиться. Она будет, как родовое проклятие, передаваться всем его потомкам вечно (если только не произойдет обратная мутация, а вероятность этого очень мала). У бесполых организмов отбор может отбраковывать только целые геномы, но не отдельные гены. Поэтому в череде поколений бесполых организмов может (при соблюдении определенных условий) происходить неуклонное накопление вредных мутаций. Одним из таких условий является достаточно большой размер генома. У круглых червей, между прочим, геномы маленькие по сравнению с другими животными. Может быть, поэтому они и могут позволить себе самооплодотворение (см. ниже). Если же организмы размножаются половым путем и практикуют перекрестное оплодотворение, то индивидуальные геномы постоянно рассыпаются и перемешиваются, а новые геномы формируются из фрагментов, ранее принадлежавших разным организмам. В результате возникает особая новая сущность, которой нет у бесполых организмов, — генофонд популяции. Гены получают возможность размножаться или выбраковываться независимо друг от друга. Ген с неудачной мутацией может быть отбракован отбором, а остальные («хорошие») гены данного родительского организма могут благополучно сохраниться в популяции. Таким образом, первая идея состоит в том, что половое размножение способствует очищению геномов от «генетического груза», то есть помогает избавляться от постоянно возникающих вредных мутаций, не допуская вырождения (снижения общей приспособленности популяции). Вторая идея сродни первой: она предполагает, что половое размножение помогает организмам эффективнее адаптироваться к меняющимся условиям за счет ускоренного накопления мутаций, полезных в данной обстановке. Допустим, у одной особи возникла одна полезная мутация, у другой — другая. Если эти организмы бесполые, у них практически нет шансов дождаться объединения обеих мутаций в одном геноме. Половое размножение дает такую возможность. Оно фактически делает все полезные мутации, возникшие в популяции, «общим достоянием». Ясно, что скорость приспособления к меняющимся условиям у организмов с половым размножением должна быть выше.
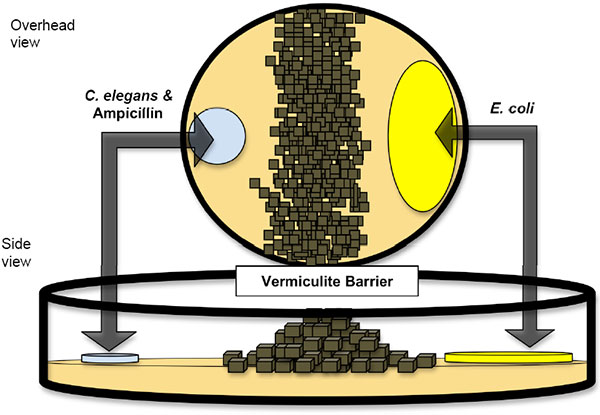
Все эти теоретические построения, однако, основаны на определенных допущениях. Результаты математического моделирования свидетельствуют о том, что степень полезности или вредности перекрестного оплодотворения по сравнению с бесполым размножением или самооплодотворением зависит от целого ряда параметров. В их числе размер популяции; скорость мутирования; размер генома; количественное распределение мутаций в зависимости от степени их вредности/полезности; число потомков, производимых одной самкой; эффективность отбора (степень зависимости числа оставляемых потомков не от случайных, а от генетических факторов) и т. д. Некоторые из этих параметров очень трудно измерить не только в природных, но и в лабораторных популяциях. Поэтому все гипотезы такого рода остро нуждаются не столько в теоретических обоснованиях и математических моделях (всего этого уже в избытке), сколько в прямой экспериментальной проверке. Однако подобных экспериментов до сих пор проведено не так уж много (Colegrave, 2002. Sex releases the speed limit on evolution // Nature. V. 420. P. 664–666; Goddard et al., 2005. Sex increases the efficacy of natural selection in experimental yeast populations // Nature. V. 434. P. 636–640). Новое исследование, выполненное биологами из Орегонского университета на круглом черве Caenorhabditis elegans, ярко проиллюстрировало действенность обоих рассмотренных механизмов, обеспечивающих преимущество тем популяциям, которые не отказываются от самцов, несмотря на их «двойную цену». Уникальный объект для изучения роли самцовЧерви Caenorhabditis elegans словно нарочно созданы для экспериментальной проверки вышеупомянутых гипотез. У этих червей нет самок. Популяции состоят из самцов и гермафродитов, причем последние численно преобладают. У гермафродитов две X-хромосомы, у самцов — только одна (система определения пола X0, как у дрозофилы). Гермафродиты производят сперматозоиды и яйцеклетки и могут размножаться без посторонней помощи путем самооплодотворения. Самцы производят только сперматозоиды и могут оплодотворять гермафродитов. В результате самооплодотворения на свет появляются только гермафродиты. При перекрестном оплодотворении половина потомства оказывается гермафродитами, половина — самцами. Обычно частота перекрестного оплодотворения в популяциях C. elegans не превышает нескольких процентов. Чтобы определить эту частоту, не обязательно наблюдать за интимной жизнью червей — достаточно знать процент самцов в популяции. Следует пояснить, что самооплодотворение — это не совсем то же самое, что бесполое (клональное) размножение, однако различия между ними быстро сходят на нет в череде самооплодотворяющихся поколений. Самооплодотворяющиеся организмы за несколько поколений становятся гомозиготными по всем локусам. После этого потомство перестает отличаться от родителей генетически, точно так же, как и при клональном размножении. У C. elegans известны мутации, влияющие на частоту перекрестного оплодотворения. Одна из них, xol-1, смертельна для самцов и фактически приводит к тому, что в популяции остаются только гермафродиты, размножающиеся путем самооплодотворения. Другая, fog-2, лишает гермафродитов способности производить сперму и фактически превращает их в самок. Популяция, в которой все особи несут эту мутацию, становится обычной раздельнополой популяцией, как у большинства животных. Авторы при помощи классических методик (путем скрещиваний, а не генной инженерии) вывели две пары пород червей с почти одинаковыми геномами, различающимися только наличием мутаций xol-1 и fog-2. Первая порода в каждой паре, с мутацией xol-1, размножается только самооплодотворением (obligate selfing, OS). Вторая, с мутацией fog-2, может размножаться только путем перекрестного оплодотворения (obligate outcrossing, OO). К каждой паре пород прилагалась третья, с таким же генетическим «бэкграундом», но лишенная обеих мутаций (wild type, WT). У пород WT частота перекрестного оплодотворения в стандартных лабораторных условиях не превышает 5%. Самцы нужны! Проверено экспериментальноС этими тройками пород были проведены две серии экспериментов. В первой серии проверялась гипотеза о том, что перекрестное оплодотворение помогает избавляться от «генетического груза». Эксперимент продолжался в течение 50 поколений (червей, разумеется, а не экспериментаторов). Каждое поколение червей подвергалось действию химического мутагена — этилметансульфоната. Это приводило к увеличению частоты мутирования примерно в четыре раза. Молодых животных помещали в чашку Петри, разделенную пополам стенкой из вермикулита (см. рисунок), причем червей сажали в одну половину чашки, а их пища — бактерии E. coli — находилась в другой половине. Червей при пересадке обрабатывали антибиотиком, чтобы очистить от случайно прилипших бактерий. В результате для того, чтобы добраться до еды, а значит получить шанс выжить и оставить потомство, черви должны были преодолеть препятствие. Тем самым экспериментаторы повысили эффективность «очищающего» отбора, который отсеивает вредные мутации. В обычных лабораторных условиях эффективность отбора очень низка, потому что черви окружены пищей со всех сторон. В такой ситуации могут выжить и размножиться даже очень слабые, перегруженные вредными мутациями животные. В новой экспериментальной установке этой уравниловке был положен конец. Чтобы переползти через стенку, червь должен быть здоров и крепок.
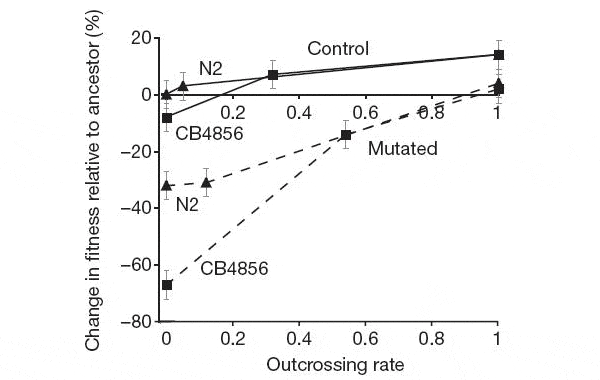
Авторы сравнивали приспособленность (fitness) у червей до и после эксперимента, то есть у особей первого и пятидесятого поколения. Червей C. elegans можно долго хранить в замороженном виде. Это сильно облегчает подобные эксперименты. Пока длился эксперимент, выборка червей 1-го поколения спокойно лежала в морозильнике. Приспособленность измеряли следующим образом. Червей смешивали в равной пропорции с контрольными червями, в геном которых был вставлен ген светящегося белка, и сажали в экспериментальную установку. Животным давали время, чтобы преодолеть барьер и размножиться, а затем определяли процент несветящихся особей в потомстве. Если этот процент увеличился в пятидесятом поколении по сравнению с первым — значит, в ходе эксперимента приспособленность выросла, если уменьшился — значит, имело место вырождение. Результаты эксперимента показаны на рисунке. Они однозначно свидетельствуют о том, что перекрестное оплодотворение является мощным средством борьбы с генетическим грузом. Чем выше частота перекрестного оплодотворения, тем лучше итоговый результат (все линии на рисунке возрастают слева направо). Искусственно повышенная скорость мутирования привела к вырождению (снижению приспособленности) всех пород червей, кроме OO — «облигатных перекрестников». Даже тем породам, у которых мутагенез не был искусственно ускорен, высокая частота перекрестного оплодотворения дала преимущество. В обычных лабораторных условиях это преимущество не проявляется, потому что червям не нужно перелезать через стенки, чтобы добраться до корма. Любопытно, что в одной из двух контрольных пород OS («облигатных самооплодотворителей») даже без повышения скорости мутирования отказ от перекрестного оплодотворения привел к вырождению (левый квадратик в верхней паре кривых на рисунке расположен ниже нуля).
Рисунок также показывает, что частота перекрестного оплодотворения у большинства «диких» пород (WT) в ходе эксперимента оказалась заметно выше исходных 5%. Это, пожалуй, самый важный результат. Он означает, что в жестких условиях (имеется в виду как необходимость перелезать через барьер, так и повышенный темп мутагенеза) естественный отбор дает явное преимущество особям, размножающимся путем перекрестного оплодотворения. Потомство таких особей оказывается более жизнеспособным, и поэтому в ходе эксперимента происходит отбор на склонность к перекрестному оплодотворению. Таким образом, первый эксперимент убедительно подтвердил гипотезу о том, что перекрестное оплодотворение помогает популяции избавляться от вредных мутаций. Во второй серии экспериментов проверялось, помогает ли перекрестное оплодотворение вырабатывать новые адаптации путем накопления полезных мутаций. На этот раз червям, чтобы добраться до пищи, нужно было преодолеть зону, заселенную патогенными бактериями Serratia. Эти бактерии, попадая в пищеварительный тракт C. elegans, вызывают у червя опасное заболевание, которое может кончится смертью. Чтобы выжить в этой ситуации, черви должны были либо научиться не глотать вредных бактерий, либо выработать устойчивость к ним. Какой из вариантов выбрали подопытные популяции червей — неизвестно, однако за 40 поколений породы OO отлично приспособились к новым условиям, породы WT приспособились несколько хуже, а породы OS не приспособились совсем (их выживаемость в среде с вредными бактериями осталась на исходном низком уровне). И снова в ходе эксперимента у пород WT под действием отбора резко возросла частота перекрестного оплодотворения. Таким образом, перекрестное оплодотворение действительно помогает популяции приспосабливаться к меняющимся условиям, в данном случае — к появлению болезнетворного микроба. Тот факт, что в ходе эксперимента у пород WT увеличивалась частота перекрестного оплодотворения, означает, что спаривание с самцами (в противовес самооплодотворению) дает гермафродитам немедленное адаптивное преимущество, которое, очевидно, перевешивает «двойную цену», которую им приходится платить, производя на свет самцов. Следует отметить, что перекрестное оплодотворение встречается не только у раздельнополых организмов. Например, многие беспозвоночные являются гермафродитами, оплодотворяющими не сами себя, а друг друга — перекрестно. У растений перекрестное опыление обоеполых («гермафродитных») особей тоже, мягко говоря, не редкость. Обе гипотезы, проверявшиеся в данной работе, вполне приложимы к таким гермафродитам. Иными словами, эта работа не доказала, что «перекрестный гермафродитизм» в чём-то уступает раздельнополости. А ведь за первый из этих двух вариантов не нужно платить пресловутую «двойную цену». Следовательно, проблема всё равно остается. Проведенные эксперименты выявили недостатки самооплодотворения по сравнению с перекрестным, но они не объяснили, почему многие организмы предпочли раздельнополость «перекрестному гермафродитизму». Ключом к решению этой загадки, скорее всего, является половой отбор. Раздельнополость дает возможность самкам придирчиво выбирать себе партнеров, и это может служить дополнительным способом повышения эффективности отбраковки вредных и накопления полезных мутаций. Возможно, и эта гипотеза когда-нибудь получит экспериментальные подтверждения. 10-10-2009 | Просмотров: 14924
Комментарии
Комментировать
Комментировать
|

Ещё по теме
|
|
|