|
|
Рыба-саламандра
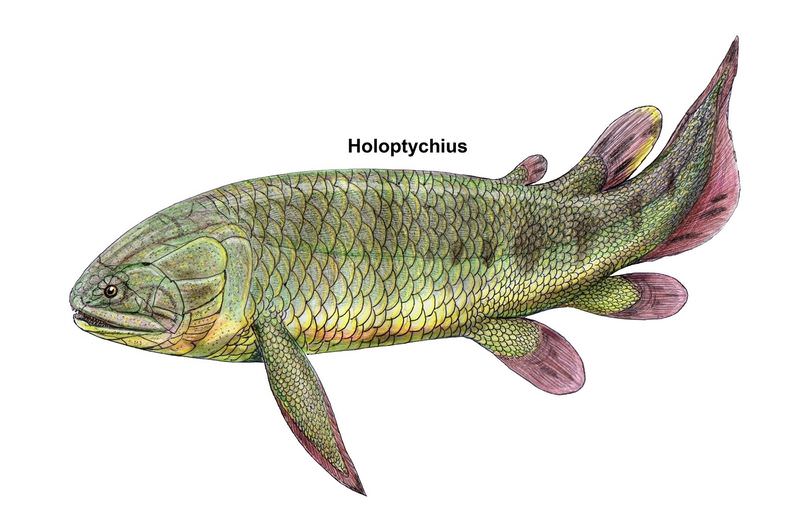
Перед нами экспонат Гётеборгского музея естественной истории (Göteborgs naturhistoriska museum): древняя кистеперая рыба голоптих (Holoptychius), которая жила в девонском периоде. На табличке рядом с экспонатом указано не только латинское название вида (Holoptychius quebecensis), но и шведское название животного — Salamanderfisk. Fisk по-шведски «рыба», значит salamanderfisk — это «рыба-саламандра». Чем голоптих заслужил такое название? Давайте разберемся, что это за существо. В широком смысле голоптих относится к группе рипидистий. Это слово требует пояснения. Название Rhipidistia ввел еще в XIX веке американский палеонтолог Эдвард Коп. В дальнейшем оно использовалось в биологической литературе очень широко. Считалось, что все кистеперые рыбы делятся на целакантов, которые всегда жили в море, и рипидистий, которые жили преимущественно (хотя и не исключительно) в пресных водах и от которых, скорее всего, произошли наземные позвоночные. Это доказывалось строением скелета, по многим признакам которого древние наземные позвоночные и рипидистии очень близки. Единственная современная кистеперая рыба — всем известная латимерия — относится к целакантам. Но нас сейчас интересуют именно рипидистии. В XX веке русский ихтиолог Лев Семенович Берг и шведский палеонтолог Эрик Ярвик (Erik Jarvik) разделили рипидистий на отряды Osteolepiformes и Porolepiformes. В результате точка зрения, принятая в учебных курсах, стала гласить, что кистеперые рыбы делятся на целакантов и рипидистий, а рипидистии, в свою очередь, делятся на остеолепиформных и поролепиформных. Проблема была в том, что рипидистии, скорее всего, не образуют единой эволюционной ветви. Сейчас, в начале XXI века, в палеонтологических работах господствует идея, что поролепиформные рипидистии эволюционно ближе к двоякодышащим рыбам, чем к остеолепиформным рипидистиям. Поролепиформов и двоякодышащих даже объединяют в группу Dipnomorpha. Это означает, что рипидистии должны либо включать в себя двоякодышащих (что начисто противоречит классической трактовке этих понятий), либо считаться сборной группой. А современная систематика тщательно избегает любых сборных групп. Поэтому в современных научных статьях, особенно англоязычных, слово «рипидистии» чаще всего не используется (даже его употребление в кавычках стало считаться дурным тоном). И если уж его использовать, то надо учитывать перечисленные оговорки.
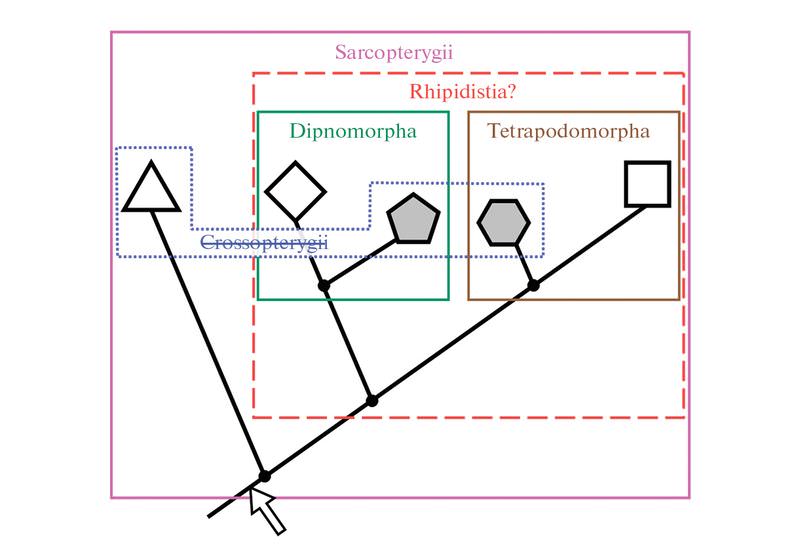 Современное эволюционное древо мясистолопастных рыб: Sarcopterygii — мясистолопастные, Crossopterigii — кистеперые, Rhipidistia — рипидистии. Обозначения: треугольник — целаканты, ромб — двоякодышащие, пятиугольник — поролепиформы, шестиугольник — остеолепиформы, квадрат — тетраподы. Вымершие группы закрашены серым. Контур группы Crossopterygii показан синим пунктиром, название группы зачеркнуто, потому что она явно сборная и в самом лучшем случае отражает эволюционный уровень, а не ветвь. Стрелка — вероятный момент образования двублочного черепа (общий признак «кистеперых»). Иллюстрация из статьи: А. Н. Кузнецов, 2018. Род Holoptychius выделил еще в 1839 году швейцарский исследователь Жан Луи Агассис, который по праву считается основателем палеонтологии рыб. Голоптих — одно из тех девонских ископаемых, что чаще всего попадаются палеонтологам. Число его находок очень велико. Обычная длина найденных голоптихов — от полуметра до метра, но есть данные, что по крайней мере некоторые виды этого рода были гораздо крупнее и достигали в длину нескольких метров (правда, от таких крупных экземпляров обычно остаются только отдельные чешуи).
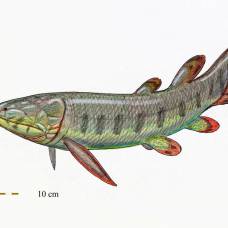
 Голоптих. Длина животного, с которого делалась эта реконструкция, — 46 см. Иллюстрация из книги E. Jarvik, 1980. Basic structure and evolution of vertebrates Как и все рипидистии, голоптих был хищником, питавшимся другими позвоночными и находившимся на вершине девонской пищевой пирамиды. Более того, он был одним из крупнейших хищников своего времени. И заодно — одним из самых успешных. Голоптихи населяли всевозможные пресные водоемы и очень широко расселились по Земле. Их остатки найдены не только в Северной Америке и Европе, но и в Южной Америке, в Колумбии, а остатки их близких родственников — даже в Австралии и Антарктиде. Возможно, что по численности и широте географического распространения голоптих занимает первое место среди всех вообще кистеперых рыб, открытых палеонтологами. По классификации Эрика Ярвика голоптих относится к отряду Porolepiformes. Характерные внешние признаки этого отряда — широкое тупое рыло и длинные грудные плавники, построенные по типу бисериального архиптеригия. Скелет такого плавника состоит из членистой оси, к каждому членику которой с двух сторон (а не с одной) прикрепляются боковые лучи. По сравнению с унисериальным архиптеригием, характерным для остеолепиформных рипидистий и для целакантов, бисериальный плавник поролепиформов является менее сильным, но более гибким. В современности этот тип плавника сохранили двоякодышащие рыбы — австралийский рогозуб, африканский протоптер и южноамериканский лепидосирен. Интересно, что если грудные плавники у голоптиха, как и положено типичным бисериальным плавникам, длинные и листовидные, то брюшные плавники — почему-то короткие и округлые. Такое сочетание уникально для поролепиформов, ни у кого больше оно не встречается.
У современных двоякодышащих, особенно у протоптера и в еще большей степени у лепидосирена, парные плавники служат не столько органами движения, сколько органами осязания — гибкими щупиками, которыми рыба исследует окружающее пространство. Парные плавники голоптиха, во всяком случае грудные, наверняка играли ту же роль. Они должны были быть гибкими и подвижными, особенно в дистальной (дальней от туловища) части. Кроме того, у голоптиха исключительно хорошо развита боковая линия, в особенности на голове. Боковая линия — это орган дистантной механорецепции, позволяющий позвоночным животным ориентироваться в воде без помощи зрения. А вот глаза у голоптиха как раз маленькие. Всё это показывает, что он искал свою добычу или в темноте, или, что гораздо вероятнее, в очень мутной воде. Из палеоэкологии известно, что в начале и середине девонского периода реки были неустойчивы, их русла постоянно мигрировали и берега размывались течением, которое, естественно, уносило с собой частицы грунта. Это и заставляло тогдашних хищников адаптироваться к охоте в почти непрозрачной мутной воде. Только к концу девона ситуация стала меняться, и этого изменения поролепиформы, как мы увидим, не пережили.
Палеонтологи пишут, что самым близким современным экологическим аналогом голоптиха, вероятно, является амазонская арапаима (Arapaima gigas). Это очень крупная хищная рыба, достигающая в длину как минимум трех метров. Она живет в сильно заросших заводях рек и ведет образ жизни не столько хищника-преследователя, сколько хищника-засадчика. Арапаима способна дышать атмосферным воздухом, но у нее для этого служат не легкие (которые предположительно были у рипидистий), а респираторный плавательный пузырь. Арапаима в целом похожа на голоптиха пропорциями тела и строением чешуи, хотя с эволюционной точки зрения эта рыба, относящаяся к отряду араванообразных, не имеет к кистеперым никакого отношения.
Отряд Porolepiformes традиционно делится на два семейства: Porolepidae и Holoptychiidae. Поролепиды, которые появились в палеонтологической летописи первыми, были многочисленны как в пресных водах, так и в морях. В отличие от них, голоптихииды населяли в основном пресные водоемы, а если выходили в море, то только в прибрежную полосу, в лагуны или эстуарии. В чисто морских отложениях остатки голоптихиид не встречаются. Это семейство специализировалась на освоении пресных вод, заняв там нишу крупных хищников. Голоптих как раз и был самым крупным представителем семейства голоптихиид. Более древние и архаичные поролепиды имели крупные ромбовидные чешуи со слоем космина — дентина, пронизанного множеством микроскопических колбовидных полостей, в которых, по некоторым предположениям, находились рецепторы каких-то органов чувств. В отличие от них, гироптихииды утратили космин и перешли на гораздо более тонкую, легкую и обтекаемую циклоидную чешую, примерно такую же, как у большинства современных рыб. Считается, что это позволяло им быстрее плавать. Сменив раннедевонских поролепид, голоптихииды в среднем девоне были вне конкуренции в роли крупнейших на Земле пресноводных хищников. Только в позднем девоне их слегка потеснили другие рипидистии — остеолепиформные. Поролепиформы — исключительно девонская группа. Они известны только из девонского периода, зато на всем его протяжении от самого начала до самого конца (от первого до последнего яруса). Верхнюю границу девона поролепиформы, насколько нам известно, не перешли. Именно в этот момент режим водоемов стал радикально меняться, и пресноводные кистеперые попали под давление, с одной стороны, быстро эволюционирующих лучеперых рыб, а с другой — четвероногих позвоночных, многие из которых вернулись в воду либо никогда ее и не покидали. Остеолепиформы смогли приспособиться к новым условиям и просуществовать в них еще какое-то время, а вот поролепиформы — не смогли. Все исследователи, работавшие с поролепиформами, отмечают их высокую эволюционную консервативность. Крупнейшим событием в истории поролепиформов было замещение поролепид голоптихиидами, но различия между этими семействами не так уж велики, если не считать типа чешуи. Внутри семейств различий еще меньше. Почти всё разнообразие, описанное внутри семейства голоптихиид, связано или с разницей в абсолютном размере (это позволяло поролепиформам разделять между собой экологические ниши: один вид — крупный хищник, другой — более мелкий), или с явно второстепенными признаками, адаптивный смысл которых неясен. По мнению некоторых исследователей, колебания таких признаков могли быть попросту случайными, вызванными процессом дрейфа генов. Никакие эволюционные тенденции (trends) внутри семейства голоптихиид не выявляются. В этом плане поролепиформные рипидистии резко контрастируют с остеолепиформными, у которых разнообразие гораздо выше и ряд тенденций отлично прослеживается, в том числе и на том же временном интервале. Причина такой разницы в темпах эволюции неизвестна. Но всё-таки почему же шведы называют голоптиха рыбой-саламандрой? Дело тут в работах шведского палеонтолога Эрика Ярвика, того самого, который разделил рипидистий на остеолепиформных и поролепиформных. Никто во всей истории мировой палеонтологии не изучал строение ископаемых кистеперых рыб так полно и детально, как Ярвик. Его любимым объектом была остеолепиформная рыба эустеноптерон (Eusthenopteron), но и поролепиформами он занимался весьма основательно. В результате у Ярвика сложилась довольно необычная точка зрения на происхождение наземных позвоночных. Он считал, что хвостатые амфибии произошли от поролепиформных рипидистий, а все остальные наземные позвоночные — от остеолепиформных. Эту гипотезу он подкрепил множеством данных, касавшихся анатомии обонятельного отдела черепа, костей нёба, верхней челюсти, жаберного и подъязычного аппарата, языка, головного мозга, нервов и сосудов головы, внутреннего уха и скелета конечностей.
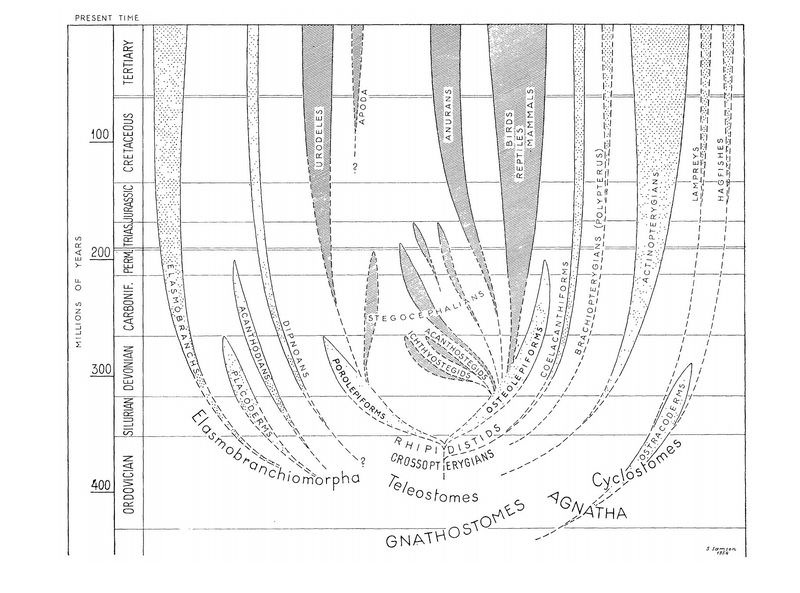 Эволюционное древо позвоночных, построенное Эриком Ярвиком. Ветви рыб и рыбообразных заточкованы, ветви тетрапод заштрихованы. Иллюстрация из статьи E. Jarvik, 1955. The oldest tetrapods and their forerunners Молекулярная филогенетика не подтвердила этот вывод Ярвика. Выяснить с ее помощью родственные отношения древних поролепиформов и остеолепиформов, разумеется, невозможно: в ископаемых образцах такой древности ни белки, ни ДНК не сохраняются. Но молекулярные родословные деревья, построенные для современных животных, показывают, что хвостатые амфибии не занимают среди них никакого особого положения. Две главные ветви, на которые распадается эволюционное древо современных наземных позвоночных, — это ветвь амфибий (бесхвостых, хвостатых и безногих) и ветвь амниот (рептилий, птиц и млекопитающих). Классические представления оказались верны. Строго говоря, такой результат не исключает, что хвостатые амфибии, бесхвостые амфибии и амниоты могут быть потомками разных кистеперых рыб. Но в таком случае трансформация кистеперых рыб в наземных позвоночных должна была бы произойти даже не дважды, а минимум трижды. Сам Ярвик вообще считал ее многократной. Он полагал, что поролепиформы и остеолепиформы уже к началу девона достигли уровня организации, который позволял им выйти на сушу ценой лишь очень небольших, чисто внешних изменений. «По своему внутреннему строению они были уже тетраподами, хотя жили в воде и выглядели как рыбы», — писал Ярвик. Если это верно, то выход кистеперых рыб на сушу мог произойти очень легко, и нет ничего удивительного, если он происходил много раз. Ярвик удивительно глубоко разобрался в сложнейшей анатомии организма древних позвоночных. Но в высказывании конкретных филогенетических гипотез, то есть предположений типа «кто от кого произошел», он, как правило, был очень осторожен. За единственным исключением: на том, что хвостатые амфибии, в отличие от всех других наземных позвоночных, произошли от поролепиформов, он категорически настаивал на протяжении сорока с лишним лет. К сожалению, молекулярная филогенетика показала, что эта гипотеза в таком виде никак не может быть верной. Даже если бы предположение о многократном выходе позвоночных на сушу подтвердилось, ветвью, самой далекой от бесхвостых амфибий, всё равно остались бы не хвостатые амфибии (как думал Ярвик), а амниоты. Тем не менее шведы, уважающие своего великого земляка, видимо продолжают считать, что в гипотезе о родстве хвостатых амфибий с поролепиформами что-то есть. Вот отсюда и «рыба-саламандра». Согласно Ярвику, голоптих — это почти хвостатая амфибия, хоть и имеющая облик рыбы. Страница, посвященная отряду поролепиформов в целом, в шведской Википедии тоже называется Salamanderfiskar, «рыбы-саламандры». Природу морфологических признаков, отличающих поролепиформов от остеолепиформов, сумела частично раскрыть Наталья Сергеевна Лебедкина. Можно сказать, что она сделала следующий шаг после Ярвика. Последний обратил внимание на то, что у поролепиформов и у остеолепиформов по-разному устроен передний отдел нёба. Скелет нёба у кистеперых рыб, как и у наземных позвоночных, состоит из нескольких покровных костей, из которых нам сейчас важны сошник и парасфеноид. Парасфеноид подстилает снизу глазничный отдел мозговой коробки, а парные сошники находятся впереди от него. У остеолепиформов сошники соединены посредине между собой, задними концами — с узким длинным парасфеноидом, а передними концами — с предчелюстными костями, которые, как и у человека, образуют передний край верхней челюсти. У поролепиформов широкий парасфеноид не заходит далеко вперед, а сошники широко расставлены и не контактируют ни с парасфеноидом, ни друг с другом. Как можно объяснить эти отличия?
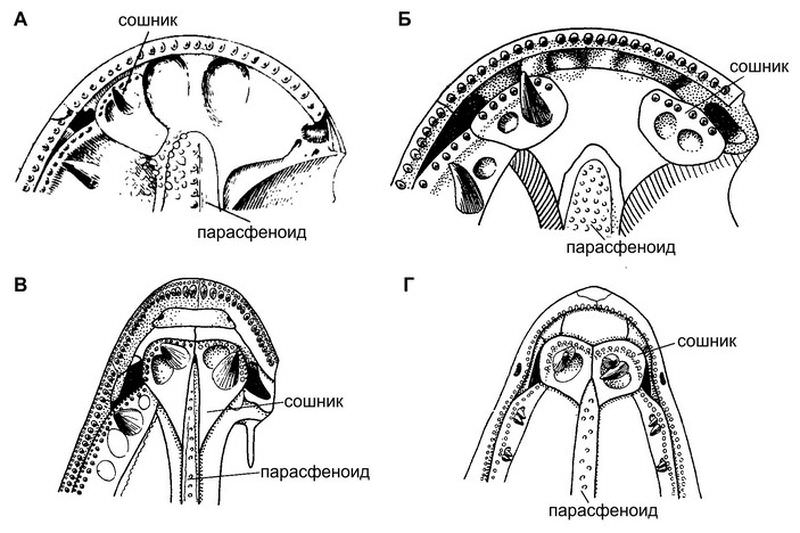 Передняя часть нёба кистеперых рыб: А — поролепиформ Poropepis, Б — примитивный остеолепофирм Thursius, В, Г — продвинутые остеолепиформы Eusthenopteron и Panderichthys. Видно, что конструкции А и Б практически не отличаются. Иллюстрации из книг: Э. И. Воробьева, 1977. Морфология и особенности эволюции кистеперых рыб и Э. И. Воробьева, 1992. Проблема происхождения наземных позвоночных Тут надо начать с самого начала. У предков рыб слизистая оболочка ротовой полости и глотки, очевидно, была покрыта многочисленными мелкими зубчиками, которые сидели на самостоятельных костных пластиночках. Однако ротовая полость и глотка позвоночных имеют еще и внутренний скелет, состоящий из висцеральных (жаберных) дуг. По мере того, как висцеральные дуги специализировались и усиливались, основания зубчиков постепенно сливались в более крупные костные пластинки. Так и возникли покровные кости нёба. Как показал тот же Ярвик, сошник и парасфеноид по своему происхождению относятся к разным висцеральным дугам: сошник — к предчелюстной дуге, а парасфеноид — к челюстной дуге (верхние элементы и той и другой у рыб вошли в состав мозговой коробки). Поэтому изначально сошник и парасфеноид не должны были образовывать никакой единой конструкции. Они ее и не образуют. Так обстоит дело не только у поролепиформов, но и у целакантов, у которых сошник вообще автономен. Однако в ходе дальнейшей эволюции костные элементы нёба стали объединяться в конструкцию, укрепляющую череп и создающую выгодное распределение механических нагрузок. Этот процесс мы видим у остеолепиформов. У продвинутых остеолепиформов он завершен: предчелюстная кость получила прочную опору через сошник на парасфеноиде, а через него — на мозговой коробке. Изящная инженерная схема. А вот что касается более примитивных остеолепиформов, то у них, как показала Эмилия Ивановна Воробьева, нёбо устроено примерно так же, как у поролепиформов: сошники широко расставлены и не контактируют ни друг с другом, ни с парасфеноидом. Это не что иное, как общий для всех кистеперых древний этап эволюции нёба. У поролепиформов он почему-то зафиксировался и сохранился до самого вымирания этого отряда. Лебедкина высказала предположение, что ключевую роль в этой истории могла сыграть предчелюстная кость, которая, как мы помним, образует передний край верхней челюсти. И у остеолепиформов, и у поролепиформов предчелюстная кость срастается с соседними покровными костями, чтобы получить надежную опору. Но происходит это по-разному. У поролепиформов предчелюстная кость срослась с костью под названием rostrale laterale, расположенной сбоку от нее. А у остеолепиформов предчелюстная кость срослась с носовой костью, расположенной сверху. Тем самым у остеолепиформов (но не у поролепиформов) возникла завершенная система продольной передачи сил: парасфеноид — сошник — предчелюстная кость — носовая кость — крыша черепа. Передняя часть мозговой коробки, расположенная как раз между носовой костью и сошником, оказалась избавлена от механических нагрузок и могла редуцироваться. По мнению Лебедкиной, это облегчило разрастание переднего мозга и тем самым — развитие интеллектуальных способностей потомков остеолепиформов, наземных позвоночных. 16-01-2025 | Просмотров: 502
Комментарии
Комментировать
Комментировать
|

Ещё по теме
|
|
|