Chorioactis - это род гриба, который содержит особь вида Chorioactis. Гриб широко известен как сигара дьявола или звезда Техаса в Соединенных Штатах, в то время как в Японии его называют кириномитаке (キ リ ノ ミ タ ケ). Этот чрезвычайно редкий гриб известен своим необычным внешним видом и непроизвольным распространением: он встречается только в избранных местах в Техасе и Японии. Тело плода, которое растет на пнях или мертвых корнях кедрового вяза (в Техасе) или мертвых дубов (в Японии), чем-то напоминает темно-коричневую или черную сигару, прежде чем расколется радиально в звездообразную структуру из четырех-семи кожистых лучей.
Внутренняя поверхность плодового тела покрыта споровой тканью, известной как гимениум, и окрашивается от белого до коричневого цвета, в зависимости от его возраста. Открытие плодового тела может сопровождаться отчетливым шипящим звуком и выпуском дымного облака спор.
Chorioactis geaster
Интересно, что, несмотря на множество мест, схожих по своим природно-климатическим параметрам с Техасом или японскими островами, эти грибы больше нигде в мире не встречаются. Ученые, проводившие генетический анализ этих двух популяций, пришли к выводу о том, что эти «грибные семьи» разделились около 19 миллионов лет назад или даже раньше.
Закрытое незрелое плодовое тело и образец, разрезанный, чтобы показать поверхность, несущую споры (hymenium).
Впервые гриб был собран в Остине, штат Техас, в 1893 году ботаником Люсьеном Маркусом Андервудом, который отправил образцы микологу Чарльзу Хортону Пеку для идентификации. В ежегодном отчете ботаника штата Нью-Йорк за этот год Пек описал этот вид как жгутик Urnula, хотя выразил сомнение по этому поводу. В 1902 году студент-миколог Элси Купфер подвергла сомнению предлагаемую классификацию различных видов в родах Urnula и Geopyxis, как это было предложено в публикации 1896 года о дискомицетах немецкого миколога Генриха Рема. Она считала, что передача вида Ремом в род Geopyxis нелогична:
«Даже внешне гриб не полностью соответствует собственному описанию Рема рода Geopyxis, в который он его помещает; текстура апотеция описывается как мясистая, стебель - короткий и иногда тонкий; в то время как у этого растения кожистый характер чаши, длина и толщина стебля являются его заметными чертами ».
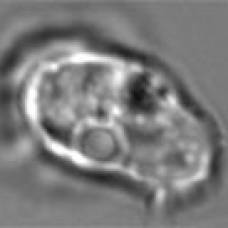
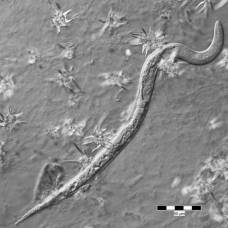
Молодые особи C. geaster имеют полое, клубовидное темно-коричневое плодовое тело, соединенное со стеблем. Стебель, который обычно закапывается в землю, короче полого плодового тела или равен его длине, хотя длина стебля несколько варьируется в зависимости от глубины подземного корня, к которому он прикреплен. Мякоть стебля и стенка плодового тела белые, а внутренняя поверхность желтовато-белая, с возрастом становится светло-коричневой. Ширина тела плода варьируется от 1,2 до 3,5 см (от 0,5 до 1,4 дюйма) в самой толстой части и имеет длину от 4 до 12 см (от 1,6 до 4,7 дюйма); ствол имеет ширину от 0,75 до 1,5 см (от 0,3 до 0,6 дюйма) и длину от 1 до 5 см (от 0,4 до 2,0 дюйма).
Образец вскоре после распада, но до расщепления на изогнутые лучи.
Как стебель, так и плодовое тело покрыты густым слоем из мягких коричневых бархатистых «волосков» или зубца. В зрелости плодовое тело распадается на 4–7 лучей, которые изгибаются вниз, подобно грибам рода Geastrum. Споры распространяются на внутренней поверхности лучей, которые, в зависимости от зрелости образца, могут варьироваться в цвете от беловатого до шафранового, от лососевого до ириски и каштанового. Кожистые лучи имеют толщину до 0,35 см (5 дюймов).
Плодовое тело остается закрытым до того, как незадолго до выброса споры; Разрушение (раскрытие тела плода) вызвано давлением, оказываемым набухшими парафизами - стерильными (то есть непроизводительными) клетками, которые вкраплены между аскоспорами. Разрушение сопровождается выпуском облаков спор, напоминающих дым.
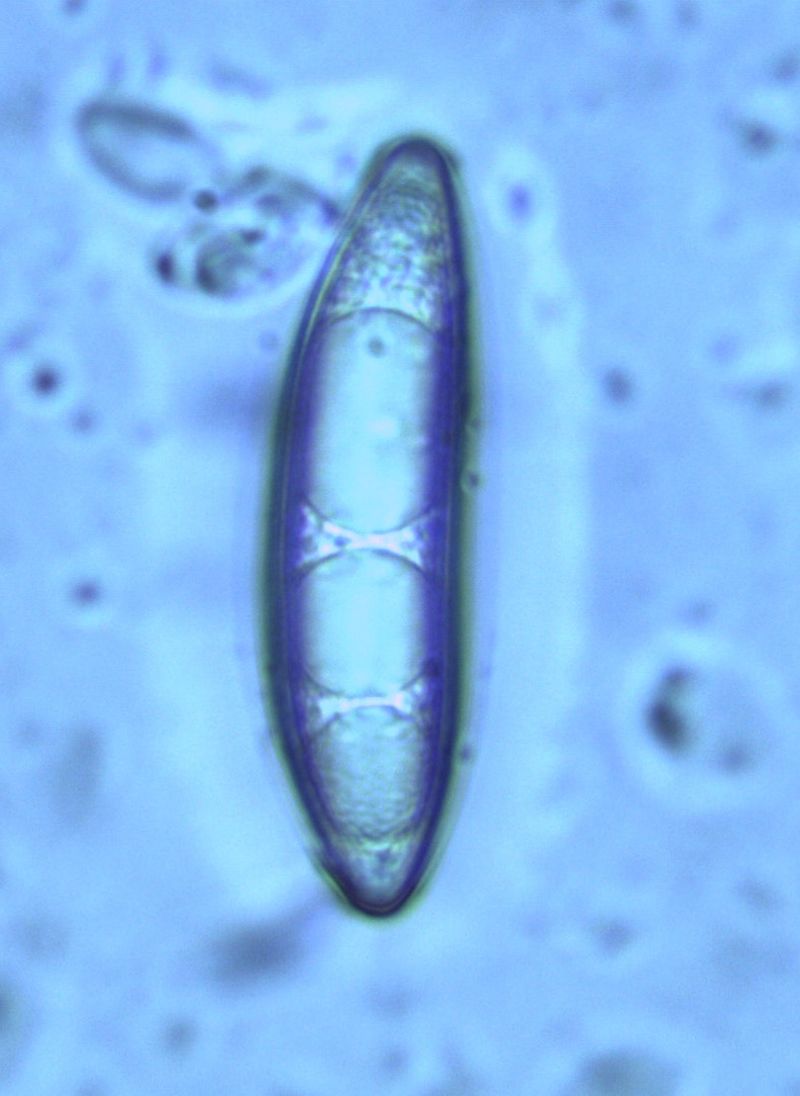
Споры имеют продолговатую или веретенообразную форму и содержат от 3 до 5 масляных капель.
Считается, что вспучивание спор при разрыве вызвано внезапным изменением относительной влажности между внутренней камерой плодового тела и внешней средой. Разрушение сопровождается шипящим звуком, явлением, которое встречается примерно у пятнадцати других видов грибов.
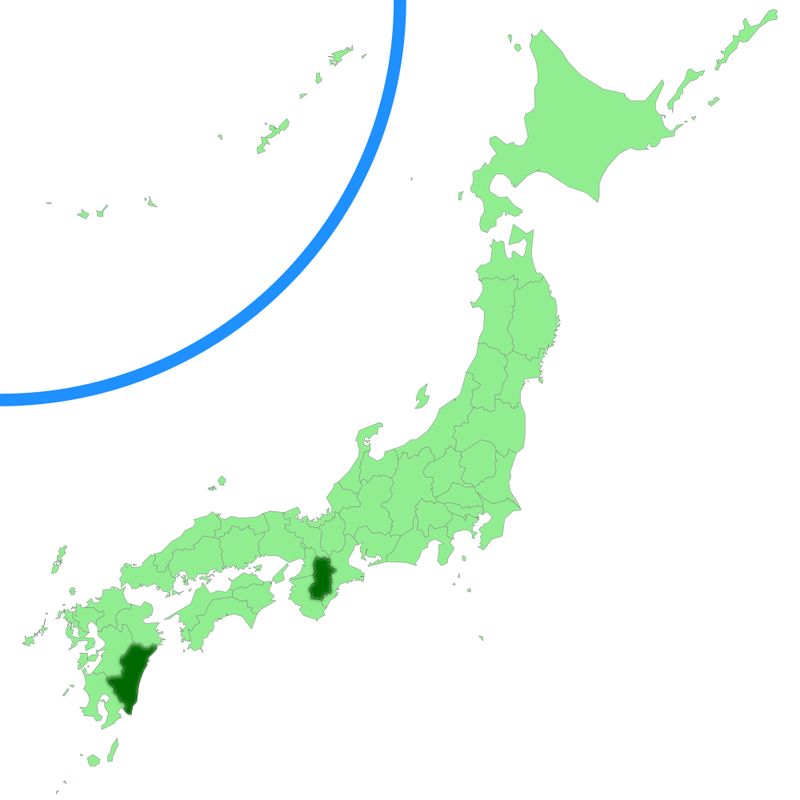
Chorioactis собирается только вТехасе и Японии. Первый зарегистрированный сбор в Японии был в Кюсю в 1937 году, а затем он снова был собран в этом месте, пока не прошло тридцать шесть лет. В 2006 году он наблюдался во влажном лесу недалеко от Каваками, префектура Нара.
Естественная среда обитания гриба в Японии исчезает из-за практики вырубки лесов и пересадки японского кедра (Cryptomeria japonica). Этот редкий гриб был включен в список исчезающих видов в Японии.
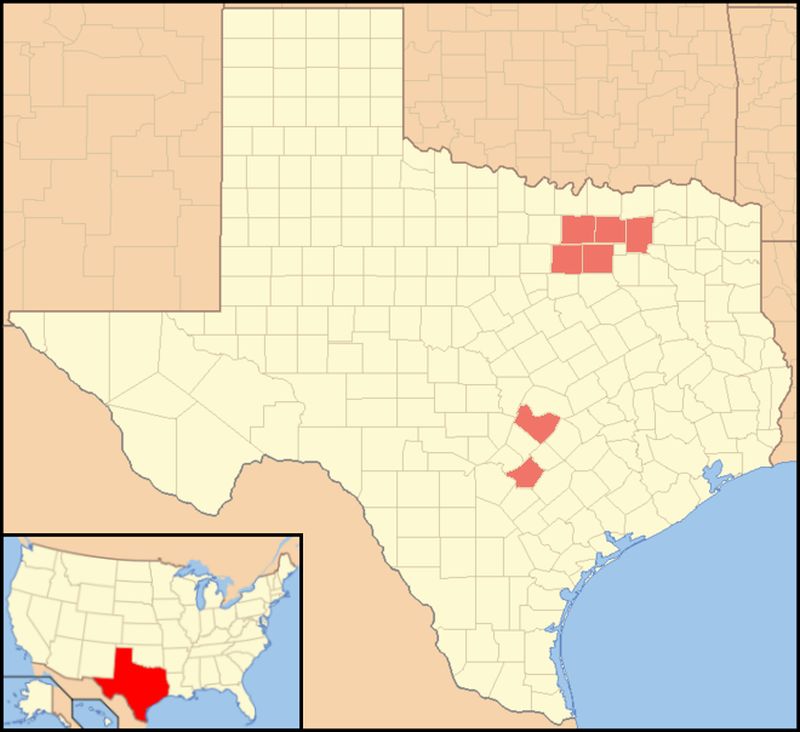
В Техасе о грибке сообщили в округах Коллин, Хейс, Тревис, Даллас, Дентон, Гваделупе, Таррант, Хант и Белл.
Хотя это не известно окончательно, Chorioactis, как полагают, является сапробным, получая питательные вещества из разлагающегося органического вещества. В Техасе плодовые тела растут поодиночке или группами из корней, пней и мертвых корней кедровых вязов (Ulmus crassifolia) или Symplocos myrtacea; в Японии обычным хозяином является мертвый дуб. Плодовые тела могут быть сгруппированы близко к основанию пня или от корней от пня; стебель плодового тела имеет тенденцию браться за точку от 5 до 10 см (2 - 4 дюйма) под землей. В Техасе плодовые тела обычно появляются между октябрем и апрелем, поскольку этот период связан с несколько более прохладной погодой, а температура и влажность в это время, по-видимому, более благоприятны для роста.
Ученые не знают, почему гриб загадочным образом живет только в Техасе и Японии, в местах примерно одинаковой широты, но разделенных на 11 000 км. Фред Джей Сивер прокомментировал: «Это всего лишь еще одна иллюстрация необычного и непредсказуемого распространения многих видов грибов. Было бы действительно трудно объяснить это, и мы просто принимаем факты такими, какие они есть». В 2004 году в исследовании сравнивались последовательности ДНК обеих популяций. Ученые пришли к выводу, что две популяции были разделены в течение, по крайней мере, девятнадцати миллионов лет, исключая возможность интродукции вида человеком из одного места в другое. Хотя между двумя популяциями нет постоянных различий в морфологии, в их жизненных историях есть несколько различий. Предпочтительным хозяином техасских популяций обычно являются корни и пни Ulmus crassifolia, в то время как японские популяции имеют тенденцию расти на упавших стволах Symplocos myrtacea и Quercus gilva. Виды техасцев растут в районах, подверженных периодическим наводнениям, в отличие от своих японских коллег. Наконец, в культуре можно выращивать только японские образцы - споры техасского материала не были успешно проращены на искусственных средах.