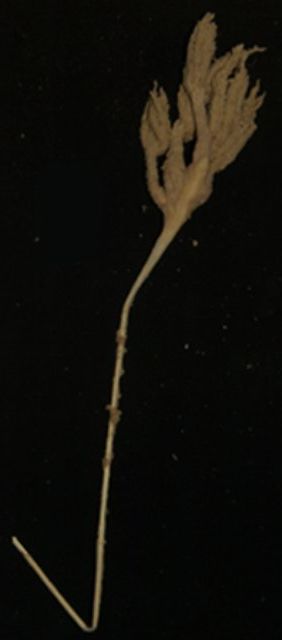
Морские перья - это потрясающие восьмилучевые кораллы. Колонии некоторых видов этих полипов по форме напоминают птичье перо.
Насчитывают около 200 видов, объединяемых в 14 семейств:
Anthoptilidae(Kölliker, 1880)
Chunellidae(Kükenthal, 1902)
Echinoptilidae(Hubrecht, 1885)
Funiculinidae (Gray, 1870)
Halipteridae(Williams, 1995)
Kophobelemnidae (Gray, 1860)
Pennatulidae(Ehrenberg, 1834)
Protoptilidae (Kölliker, 1872)
Renillidae (Gray, 1870)
Scleroptilidae (Jungersen, 1904)
Stachyptilidae (Kölliker, 1880)
Umbellulidae(Kölliker, 1880)
Veretillidae (Herklots, 1858)
Virgulariidae (Verrill, 1868)
Большинство из 200 современных видов обитают на больших глубинах, лишь немногие распространены в прибрежных водах.
Из всех кишечнополостных самые фантастические по форме колонии образуют морские перья, относящиеся к восьмилучевым кораллам:
Виргулярия (Virgularia)
Veretillum cynomorium
Умбеллула (Umbellula)
Кофобелемнон (Kophobelemnon stelliferum)
В отличие от других колониальных кишечнополостных морские перья никогда не прикрепляются к какому-нибудь субстрату. Их нижний конец, имеющий обычно мускулистое расширение, свободно втыкается в мягкий грунт, выстилающий дно. Хотя морские перья ведут неподвижный образ жизни, они могут в случае необходимости передвигаться по грунту и вновь укрепляться на новом месте.
Морские перья образуют неразветвленные, часто довольно крупные и ярко окрашенные колонии. Они состоят из мясистого ствола, представляющего собой разросшееся и сильно видоизмененное тело первичного полипа. Остальные полипы вырастают на стенках ствола или на его выростах. Для всех морских перьев характерен диморфизм вторичных полипов. Имеются крупные кормящие полипы — аутозоиды — и мелкие недоразвитые полипы — сифонозоиды. Каждая колония морского пера делится на два отдела: верхний, или полипоидный, и нижний — стебель. На полипоидной части колонии образуются аутозоиды и сифонозоиды. Стебель свободен от полипов, хотя у некоторых видов и здесь развиваются мелкие сифонозоиды.
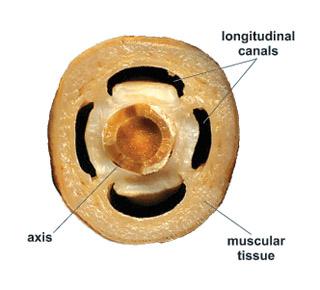
Для морских перьев характерна высокоорганизованная система каналов, связывающая различные отделы колонии. Она состоит из центральной и периферической систем. Центральная система представлена четырьмя основными продольными каналами, пронизывающими всю колонию. Два из них являются продолжением радиальных карманов гастральной полости первичного полипа. Два других канала — вторичные образования. Периферическая система состоит из сложной густой сети каналов, связывающих между собой гастральные полости отдельных полипов. Каналы периферической системы открываются в основные продольные каналы колонии.
Umbellula encrinus.
Скелет морских перьев слагается из многочисленных известковых спикул в виде палочек и игл, обычно имеющих три продольных ребрышка. Кроме спикул, образующих, как правило, корковый слой тела колонии, имеется и осевой скелет, образующийся, так же как и у роговых кораллов, из концентрических роговых пластинок, пропитанных известью.
Морским перьям свойственна хорошо развитая мышечная система. В ней различают мощную кольцевую мускулатуру, отражающую продольные каналы, слой продольных мышечных тяжей и слой косых мускулов, прикрепляющихся одним концом к стержню осевого скелета. Косые мускулы обеспечивают произвольное изгибание колонии.
Нервная система морских перьев достигает высокого для кишечнополостных совершенства. Основная черта ее прогресса заключается в образовании общеколониальной нервной системы, связывающей между собой все отделы колонии. Однако специальные органы чувств отсутствуют и у морских перьев.
Питание пеннатулярий изучено еще недостаточно. В гастральных полостях аутозоидов Sclerobelemnon были обнаружены мелкие многощетинковые черви из семейства Syllidae. Имеются указания на то, что пищей для Protoptilum служат крылоногие моллюски, а для Virgularia — личинки различных двустворчатых моллюсков. Возможно, что морские перья питаются и мелкими взвешенными частицами органического вещества животного.
Морские перья раздельнополы. Как правило, женские колонии оказываются гораздо более многочисленными, чем мужские. Половые клетки образуются, в отличие от Alcyonaria и Gorgonaria, обладающих диморфизмом полипов, исключительно в аутозоидах. Сифонозоиды морских перьев обеспечивают лишь циркуляцию воды и, следовательно, газообмен колонии. В большинстве случаев оплодотворение зрелых яиц пеннатулярий происходит в наружной среде. Зрелые яйца попадают сначала в гастральную полость, а затем антиперистальтическими сокращениями глотки выбрасываются в воду. Яйца у многих видов пеннатулярий выбрасываются ранним утром, независимо от температуры воды. Нерест колоний происходит одновременно, при этом мужские колонии выбрасывают половые продукты в таком количестве, что вода вокруг них становится молочно-белой. У северных видов пеннатулярий нерест может продолжаться весь летний период.
Лишь немногие виды пеннатулярий можно считать живородящими. У таких видов оплодотворение происходит в гастральной полости, а в воду выпускаются личинки на стадии планулы.
Как правило, почкование приводит у морских перьев, так же как и у других колониальных полипов, лишь к росту колоний, ибо новообразованные полипы не теряют связи с материнским организмом. Только у одного вида — Pennatula phosphorea — известно размножение делением, при котором сразу образуется новая колония. Происходит это следующим образом. Верхняя часть молодой колонии, на которой имеется несколько полипов и куда не доходит осевой стержень, начинает отшнуровываться и в конце концов полностью отделяется от материнской колонии. У отделившейся части образуется вначале стебель, при помощи которого новообразованная колония укрепляется в грунте, а затем и осевой стержень.
Морские перья распространены всесветно. Однако в приантарктических водах, за исключением трех видов умбеллул, морские перья не встречаются. Большинство видов обитает в тропическом и умеренном поясах северного полушария. В северных частях Атлантического и Тихого океанов число видов пеннатулярий резко снижается, а в холодных водах полярного бассейна встречается только три вида — Umbellula encrinus, Virgularia cladiscus и V. glacialis. На мелководье обитает 42% всех известных видов пеннатулярий. Однако распределение подавляющего большинства мелководных видов, относящихся всего лишь к нескольким родам, ограничено теплыми водами тропической зоны. В нижней сублиторали и в батиали обитает более половины всех видов пеннатулярий. На глубину более 1000 м проникает 32 вида, а на глубину более 3000 м — 16, или 7% всех известных пеннатулярий. Некоторые глубоководные пеннатулярий представляют собой весьма характерный элемент абиссальной донной фауны. Завоевать большие глубины позволили им необычайная экологическая пластичность, а также то, что морские перья приспособились к обитанию на мягких илистых грунтах, которые, естественно, и преобладают на больших глубинах.
Несколько видов пеннатулярий способны переносить значительное опреснение воды. Один из таких видов — Virgularia mirabilis, широко распространенный в Северной Атлантике и Средиземном море, смог проникнуть поэтому даже в Черное море. Другой вид — Renilla patula — был найден в устье реки Миссисипи.
Veretillum cynomurium
Рассмотрим несколько наиболее характерных представителей отряда. В Атлантическом океане, вдоль берегов Европы и Африки от Бискайского залива до Анголы, а также в Средиземном море на глубине 20—40 м часто встречается Veretillum cynomurium. Этот вид принадлежит к наиболее просто организованным морским перьям. Колония веретиллума, достигающая 40 см в длину, имеет цилиндрическую форму. На поверхности полипоидной ее части, составляющей три четверти общей длины колонии, беспорядочно разбросаны десятки крупных, до 7, 5 мм, аутозоидов с длинными щупальцами. Сифонозоиды еще более многочисленны, но они расположены в продольных рядах. Осевой скелет редуцирован до гладкой палочки длиной в 10—15 мм, а у некоторых экземпляров вообще отсутствует. Значительно редуцированы испикулы. Они очень мелки и имеют овальную форму.
Веретиллум — излюбленный объект для наблюдения в аквариуме. В состоянии покоя колонии этого морского пера выглядят довольно невзрачно: они имеют вид коротких желто-коричневых палочек, косо торчащих в грунте. Однако ночью веретиллум преображается. Колония всасывает воду, расправляется и превращается в почти прозрачный желто-оранжевый высокий цилиндр, на стенках которого распускаются прозрачные, совершенно белые полипы. Через некоторое время распустившаяся колония начинает фосфоресцировать. При незначительном раздражении колонии свечение усиливается, и по всей колонии пробегают волны света. К утру свечение прекращается, полипы втягиваются, а сама колония сжимается и приобретает свой обычный вид сморщенной палочки.
Наличие общеколониальной нервной системы обеспечивает быструю и согласованную реакцию всего организма на механические или электрохимические раздражения, приложенные к различным отделам колонии. Достаточно 20—30 секунд после нанесения раздражения, чтобы вся колония веретиллума оказалась в сокращенном состоянии.
К числу примитивных морских перьев относятся виды из семействa Kophobelemnonidae, положение аутозоидов которых не совсем упорядоченно. Однако их колониям уже присуща двусторонняя симметрия — признак некоторого усовершенствования колоний. Все представители семейства имеют уже хорошо развитый осевой скелет, а известковые спикулы многих видов приобретают обычную для остальных пеннатулярий форму длинных палочек с тремя продольными ребрышками. Многие виды рода Kophobelemnon приспособились к обитанию на больших глубинах океанов. По мере завоевания глубин происходило и значительное видоизменение колоний кофобелемнон. Если наиболее мелководные представители рода обладают несколькими десятками мелких полипов, рядами расположенных на полипоидной части колонии, то у наиболее глубоководных видов — Kophobelemnon pauciflorum и Kophobelemnon biflorum, обитающих в абиссали Индийского и Тихого океанов, образуется соответственно только три и два крупных аутозоида. Уменьшение числа аутозоидов при переходе к глубоководному образу жизни — одно из проявлений приспособительной олигомеризации, о которой говорилось выше, в разделе, касающемся общего строения колоний кишечнополостных.
Umbellula encrinus
Umbellula carpenteri
Umbellula thomsoni.
Более высокой организации достигли колонии умбеллул — чрезвычайно своеобразных морских перьев. Колония умбеллулы состоит из необычайно длинного и тонкого стебля, на вершине которого помещается чашевидный рахис (так называется у морских перьев полипоидный отдел колонии). Его верхняя поверхность несет несколько очень крупных, не втягивающихся аутозоидов с длинными щупальцами. Многочисленные сифонозоиды почти сплошь покрывают нижнюю поверхность рахиса и пространство между аутозоидами. Любопытно, что они имеются на стволе колонии и даже на мясистом его расширении, при помощи которого колония укрепляется в грунте. В целом колония умбеллулы напоминает цветок амариллиса. Большинство видов умбеллул — обитатели холодноводных районов и больших глубин океана. Только эти морские перья населяют антарктические воды южного полушария (Umbellula еncrinus, U. magniflora и U. thomsoni). U. encrinus, кроме того, широко распространены в Северном Ледовитом океане (рис. 176). Рекордная глубина обитания восьмилучевых кораллов — 6120 м — также принадлежит умбеллуле U. thomsoni. Представители рода Umbellula, так же как и Kophobelemnon, дают яркие примеры приспособительной олигомеризации, связанной с переходом к глубоководному образу жизни. Так, на рахисе нового вида U. monocephalus, найденного недавно в Индийском океане на глубине свыше 3000 м, имеется всего лишь один аутозоид необычайно крупного размера.
Наиболее мелкие виды умбеллул достигают 40—60 см, а более крупные, такие, как U. encrinus или U. magniflora, — даже 200 см в длину. Большой длиной стебля (179 см) обладает и U. monocephalus. Размеры аутозоидов умбеллул, как правило, не превышают 2—3 см в длину. Единственный тринадцатисантиметровый аутозоиди. monocephalus кажется исполином по сравнению с обычными аутозоидами умбеллул.
Однако высшим типом организации обладают те морские перья, у которых аутозоиды отходят не непосредственно от боковой поверхности главной оси, а от особых ее выростов, называемых листьями. Именно такие колонии по внешнему виду наиболее сравнимы с пером птицы. Это сходство и привело к возникновению названия рассматриваемой группы кишечнополостных .
К морским перьям, аутозоиды которых сидят на боковых листьях колонии, относится очень изящный вид Scitaliopsis djiboutiensis, обитающий в Индийском океане у берегов Сомали. Наблюдения над тем, как вытащенное из грунта морское перо вновь укрепляется в нем, производились именно над этим мелководным видом. Положенная на грунт, колония некоторое время оставалась совершенно неподвижной. Затем нижний конец «стебля» начал медленно изгибаться подобно геотропическому изгибу корня растения. При этом по нему стала распространяться волна мышечных сокращений. То он вытягивался и становился очень тонким, то расширялся и сокращался в длину. При этом нижний конец «стебля» колонпп постепенно начал вклиниваться в грунт.
По мере того как стебель все глубже и глубже проникал в грунт, колония все больше и больше приподнималась над ним, пока не приняла свое обычное вертикальное положение.
Чарлз Дарвин во время своего путешествия на корабле «Бигль» наблюдал поведение морских перьев Stylatula darvinii, образующих плотные поселения на мелководье залива Байя Бланка. Он писал, что там во время отлива можно видеть сотни этих «зоофитов», неподвижно торчащих над поверхностью грунта. Но достаточно только прикоснуться к такой имеющей несколько дюймов в длину колонии, как она с силой начинает вбуравливаться в ил и быстро в нем исчезает.
Очень красивые колонии образует морское перо Pennatula phosphorea. Этот вид чрезвычайно широко географически распространен. В Атлантическом океане он встречается от Гренландии до субантарктических вод, в Тихом океане — от Берингова моря до берегов Японии. Колонии P. phosphorea были найдены и в Индийском океане. Это морское перо обладает широким диапазоном вертикального распределения, обитая как на мелководье, так и в абиссальных глубинах океана. Мелководные экземпляры Р. phosphorea достигают 30 см в длину и состоят из мясистой главной оси, от которой в полипоидной части отходят на близком расстоянии друг от друга многочисленные узкие треугольные «листья». По краям последних сидят мелкие аутозоиды, количество которых определяется длиной листа. На самых длинных листьях насчитывается до 25 полипов. Каждый аутозоид защищен чашечкой из длинных палочковидных спикул, заканчивающейся несколькими острыми зубцами. Глубоководные экземпляры не столь красивы. Они гораздо мельче, листья их более редки. На верхнем крае листа, скорее имеющего вид тонкой веточки, располагается всего 3—4 аутозоида.
Pennatula phosphorea
Колонии P. phosphorea обычно окрашены в ярко-красный цвет, хотя встречаются экземпляры с более светлой окраской— оранжево-красные, желтые и почти белые. Своей окраской колонии P. phosphorea обязаны спикулам, ибо только они окрашены в красный или желтый цвет. Мягкие же ткани совершенно бесцветны. Белые полипы распустившейся колонии красиво контрастируют с яркой окраской листьев.
Virgularia mirabilis и P. phosphorea.
В пассивном состоянии пеннатула почти плашмя лежит на грунте, но в активном состоянии она начинает быстро впитывать воду и принимает обычное для морских перьев вертикальное положение. Поникшие листья ее при этом расправляются. Как и у других мелководных форм, у P. phosphorea существует правильное чередование в течение суток активного и пассивного состояния. В активном состоянии колонии пеннатулы находятся в ночное время.
P. phosphorea
Подобно веретиллумам и многим другим пеннатуляриям, колонии пеннатулы способны к свечению. Свет излучается особой слизью, выделяемой при раздражении железистыми эктодермальными клетками. У некоторых видов светятся все отделы колонии, у других — только полипоидная часть, у третьих — только полипы. У веретиллум, в частности, способна излучать свет поверхность колонии, у пеннатулы фосфоресцируют только аутозоиды и сифонозоиды. Если некоторые виды излучают свет все время, как, видимо, многие глубоководные морские перья, то другие светятся или только в ночное время, или в условиях полной темноты.
В дневное время такие животные начинают светиться в ответ на какое-нибудь раздражение — механическое, электрическое или химическое. Слабое раздражение вызывает вспышку света лишь в том месте, к которому был приложен раздражитель. Но если оно достаточно сильно, то по всей колонии начинают пробегать концентрические «волны» световых потоков. Световая «волна» может распространяться по колонии со скоростью 5 см в секунду. Отсеченные от колонии части тела также начинают светиться в ответ на их раздражение. Любопытно, что даже высушенная слизь морских перьев не теряет способности к люминесценции. Чтобы вызвать свечение, достаточно слегка смочить высушенную слизь водой. Экспериментально доказано, что для вызывания и поддерживания состояния люминесценции необходим постоянный приток воды, насыщенной в достаточной степени кислородом. У большинства видов пеннатулярип излучается синий или фиолетовый поток света, но у некоторых видов он может быть желтоватым и даже зеленым.