|
|
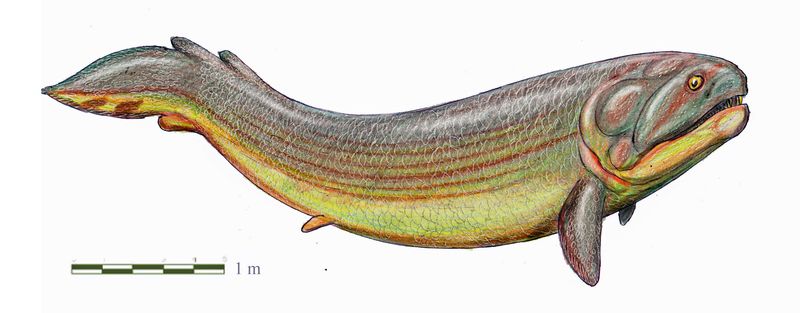
Ризодус (лат. Rhizodus)
Это драконоподобное существо называется ризодус (Rhizodus). Свое название, буквально означающее «корнезуб», оно получило в XIX веке от великого Ричарда Оуэна. Ризодус — рыба, принадлежащая к древней и могучей, хотя и почти вымершей в наши дни, эволюционной ветви мясистолопастных (саркоптеригий). Разумеется, это не наш современник. Ризодус жил в Европе в карбоне, в интервале между датами (округленно) 330 и 300 миллионов лет назад, и достигал огромного размера — до 7 метров. Прежде чем рассуждать про эту замечательную рыбу, взглянем на те ее особенности, которые хорошо видны прямо на картинке. Начнем с плавников. Как известно, рыбьи плавники можно разделить на парные и непарные. Парные плавники — грудные и брюшные, непарные — хвостовой, спинные (у мясистолопастных рыб их два) и анальный. Итак, первым делом мы видим, что грудные плавники у ризодуса очень мощные. На таких плавниках можно перемещаться по мелководью и при необходимости даже по суше. Но полноценная ходьба с их помощью, конечно, невозможна, тем более что брюшные плавники у ризодуса маленькие. Для чего его парные плавники подходят отлично, так это для опоры передней части тела в случаях, когда надо либо сидеть в засаде, либо высунуть из воды голову. О непарных плавниках обычно говорят меньше, но они тоже важны для рыбьего плавания. Рыба, у которой нет спинных и анальных плавников, теряет в маневренности, как стрела, лишенная оперения. У ризодуса эти плавники есть. Но они, во-первых, очень невелики, и во-вторых, сильно смещены назад. Второй спинной и анальный плавники, расположенные на одном уровне, отодвинуты назад буквально до упора — до самого хвоста, так что функционально они вошли в состав хвостовой лопасти. Да и первый спинной отодвинут назад так сильно, что наехал на второй. Очевидно, для ризодуса такое решение оказалось более выгодным, чем редуцировать эти плавники полностью (как произошло у пандерихта, о котором мы еще упомянем). Но хорошим пловцом он точно не был. Наконец, хвостовой плавник. У ризодуса он не расширенный, как у большинства рыб, а скромный, клиновидный, напоминающий хвост какого-нибудь тритона, только гораздо более короткий. Такой хвост называется дифицеркальным. Хвостовой плавник, как правило, служит у рыб главным движителем при активном плавании. У хороших пловцов он никогда не редуцируется, но ризодус, как мы уже выяснили, к хорошим пловцам не относился. Он был подстерегающим хищником, делающим ставку не на преследование добычи, а на короткий и мощный рывок из засады. Голова у ризодуса довольно большая, пасть, соответственно, тоже. По краям пасти сидят длинные острые клыкообразные зубы. Основания этих зубов были погружены в челюсть, примерно как у крокодилов, причем довольно глубоко (пишут, что чуть ли не на половину общей длины зуба). Отсюда и «корнезуб». Несомненно, это укрепление зубного аппарата, служащее адаптацией к захвату крупной и сильной добычи. Ризодус не уникален: он член целого семейства, которое называется Rhizodontidae. Другие представители этого семейства похожи на ризодуса, и для них тоже характерен огромный размер. Ризодус — самый крупный из них (насколько пока известно), но и для других родов сообщения о длине в пять или шесть метров — не редкость. Возможно, это крупнейшие пресноводные рыбы всех времен. Правда, иногда пишут, что современная белуга бывает аж девятиметровой, но, во-первых, это непроверенные данные (надежно описанные особи достигали лишь четырех с небольшим метров), а во-вторых, белуга — не пресноводная рыба. Она в основном живет в морях, а в реки заходит только для нереста. Так что рекорд ризодонтид она не побивает. В свою очередь, ризодонтиды принадлежат к группе, которую по традиции называют кистеперыми рыбами (кроссоптеригиями). Будем помнить, что кистеперые рыбы — не таксон. Это эволюционный уровень, объединяющий всех мясистолопастных рыб, кроме двоякодышащих (дипной) и тетрапод, то есть позвоночных с лапами. По сравнению с типичными кистеперыми и дипнои, и тетраподы — существа очень сильно специализированные, хотя и в совершенно разных направлениях. Но кистеперые рыбы интересны отнюдь не только в качестве предков этих групп. Кистеперые — это целый мир, состоящий из множества эволюционных ветвей и жизненных форм, который просуществовал несколько десятков миллионов лет (даже если не учитывать доживших до нашего времени целакантов) и обладал собственной сложной историей. Какое же место в этой истории занимают ризодонтиды? Чтобы понять это, вернемся к самому началу — к условиям, в которых кистеперые рыбы когда-то сформировались.
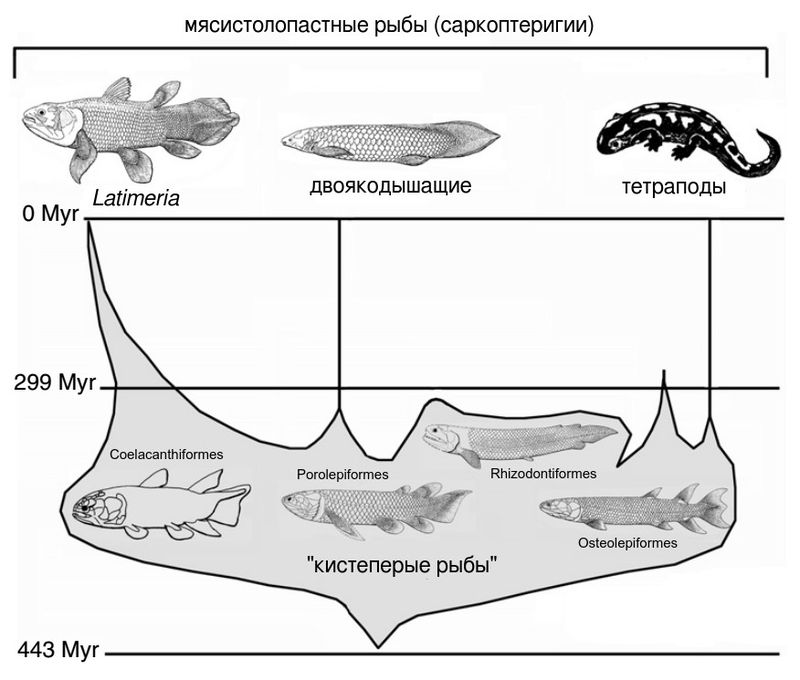 Место кистеперых рыб на эволюционном древе и место ризодонтид среди кистеперых. Потомки кистеперых — двоякодышащие рыбы и тетраподы — к ним уже не относятся. Слева мы видим ствол целакантов (Coelacanthiformes), которые наиболее обособлены от всех прочих мясистолопастных и, единственные из кистеперых, дожили до нашего времени. Они нас сейчас не интересуют. Гораздо более близкими к наземным позвоночным традиционно считались полностью вымершие поролепиформные (Porolepiformes) и остеолепиформные (Osteolepiformes) кистеперые. Причем у остеолепиформов признаков сходства с тетраподами гораздо больше. Великий шведский палеонтолог Эрик Ярвик (Erik Jarvik) считал, что хвостатые амфибии произошли от поролепиформных кистеперых, а все остальные тетраподы — от остеолепиформных. Но в наше время эта гипотеза не поддерживается. Чаще полагают, что тетраподы — потомки остеолепиформов, а поролепиформы гораздо ближе к двоякодышащим рыбам, как, собственно, и показано на рисунке. И наконец, где-то между поролепиформами и остеолепиформами, вероятно ближе к последним, отходит еще один ствол — ризодонтиформные кистеперые (Rhizodontiformes), представленные семейством Rhizodontidae. Эта группа никаких потомков не оставила, зато ее собственная эволюция была очень яркой. Вот о ней мы сейчас и говорим. Иллюстрация из работы P. Janvier, 2007. Living primitive fishes and fishes from deep time (с изменениями)
Типичным местообитанием ранних кистеперых рыб были континентальные водоемы начала девона, отличавшиеся крайне нестабильным гидрологическим режимом. Реки, текущие в неустойчивых берегах, постоянно размывали их, меняя русла и заставляя рыб либо следовать за уходящей водой, либо зарываться в ил и впадать в спячку (как это и сейчас делают африканские двоякодышащие). Кистеперые, судя по всему, предпочитали первый путь. Американский палеонтолог Альфред Ромер (Alfred Romer) считал, что они жили в водоемах, которые регулярно пересыхали, и были поэтому вынуждены в случае засухи переползать по суше в более постоянные водоемы. Чтобы облегчить ползанье, у некоторых рыб плавники превратились в лапы. Именно так Ромер объяснял появление наземных позвоночных. В XX веке этот сценарий стал очень популярным и вошел в учебники. Однако против него быстро появились возражения. Зоологи, занимавшиеся современными амфибиями, отметили, что они достаточно часто живут в пересыхающих водоемах, но такого поведения, какое предполагал Ромер для кистеперых рыб, у них никогда не наблюдается. Если водоем пересыхает, амфибии зарываются поглубже в ил и впадают в летнюю спячку, но никуда не мигрируют. Нет оснований предполагать, что предки амфибий в той же ситуации вели себя иначе. К тому же места, где жили раннедевонские рыбы, совсем не обязательно были засушливыми: девонские континенты не были сплошной пустыней, а если и были, то не столько сухой пустыней, сколько влажной. Без наземной растительности, которая могла бы образовать сплошной покров, пустыня сохранялась даже под регулярными тропическими дождями. Нам сейчас такие ландшафты незнакомы, но в девоне лик Земли был иным. Девонские реки были мелкими, кислорода в воде там было мало. Поэтому у большинства (а может, и у всех) рыб, которые там жили, появились легкие. Пресноводные кистеперые, как и двоякодышащие, сочетали жаберное дыхание с легочным. С небольшой глубины им было нетрудно подняться к поверхности, чтобы глотнуть воздуха. Кроме того, граница между водой и сушей тогда была предельно размыта. Береговой линии не существовало, огромные пространства скорее всего занимали протяженные заросшие мелководья, напоминающие современные марши. Естественно, что кистеперые рыбы, уже имевшие легкие, стали осваивать эти мелководья, тем самым переходя к более или менее амфибиотическому образу жизни. Начался процесс тетраподизации — постепенного приобретения рыбами облика тетрапод.
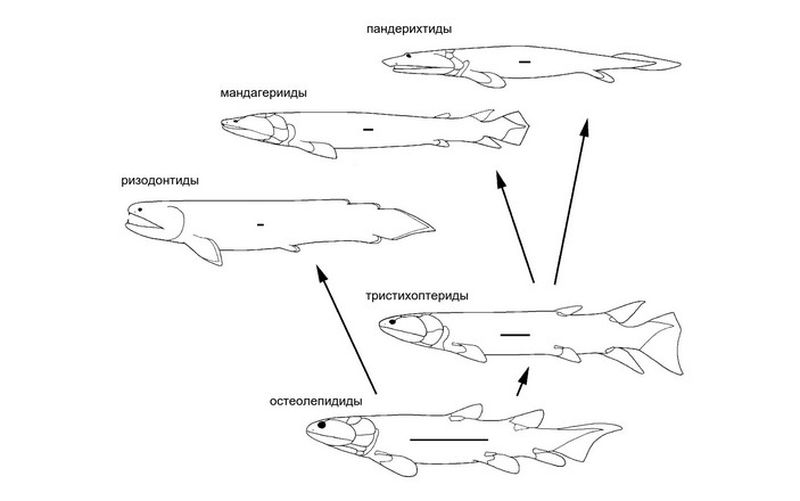 Параллельная тетраподизация кистеперых рыб. Главные пояснения — в тексте. Замечание для тех, кто внимательно читал подпись к предыдущей картинке: здесь показаны только остеолепиформные и ризодонтиформные кистеперые, но не поролепиформные и не целаканты. В общем, речь идет о ветвях, близких к тетраподам. Сами тетраподы, если бы они были на этом рисунке, находились бы рядом с пандерихтидами. Мандагериевые рыбы выделены в особую группу (в цитируемой статье этого нет, но сейчас они считаются как минимум самостоятельным подсемейством). Внутри каждой рыбы нарисована масштабная линейка длиной примерно 5 сантиметров, поэтому видно, что тетраподизация означает в том числе и увеличение размеров. Заодно можно видеть, что мандагерииды и пандерихтиды приобретают в целом вполне амфибийный облик, если не считать отсутствия тетраподных лап. О ризодонтидах этого сказать нельзя. Их облик, как и путь эволюции, — очень своеобразный. Так всегда обстоит дело при параллельной эволюции: несколько ветвей вписываются в общую тенденцию, но при этом, присмотревшись, можно увидеть, что в каждой из них сочетание признаков уникально. Например, между мандагериидами и пандерихтидами тоже есть отличия, просто они не столь заметны на картинке. Реальности самой тенденции это, разумеется, не отменяет. Иллюстрация из статьи P. Ahlberg, Z. Johanson, 1998. Osteolepiforms and the ancestry of tetrapods (с изменениями)
В 1998 году Пер Альберг (Per E. Ahlberg) и Зерина Джохансон (Zerina Johanson) опубликовали статью, в которой показали, что тетраподоподобный комплекс признаков возникал в разных ветвях кистеперых рыб минимум трижды. Очевидно, всё началось с некрупных кистеперых (длиной от 20 до 70 сантиметров), которые были относительно неспециализированы и ни о какой суше еще не помышляли. Хорошим примером такой рыбы может послужить остеолепис. Но затем в некоторых ветвях рыб начинает трансформироваться целая группа признаков, на первый взгляд не связанных между собой. Во-первых, увеличивается размер животных. Во-вторых, редуцируются непарные плавники: хвостовой плавник превращается из расширенного («рыбьего») в клиновидный («тритоний»), а спинные и анальный плавники могут вовсе исчезнуть. Парные плавники, особенно грудные, при этом, наоборот, укрепляются. В-третьих, перестраивается зубная система. Все кистеперые рыбы — хищники. Большинство хищных рыб питается другими рыбами, желательно такого размера, чтобы их можно было глотать целиком. Главная забота хищника — не дать скользкой добыче выскользнуть в момент захвата, а для этого ее надо сразу наколоть на острые «клыки», которые могут находиться и в глубине рта, на нёбе (так даже выгодней). Ротовой аппарат обычных кистеперых рыб, вроде остеолеписа, подстроен под эти задачи. Но если вы либо охотитесь не только в воде, либо интересуетесь очень крупной добычей, от которой придется отрывать куски, то вам такой ротовой аппарат не подойдет. Тогда нужно перенести «клыки» на внешние края рта, создавая универсальный живой капкан. Нетрудно видеть, что у ризодонтид наблюдаются все только что перечисленные признаки. Это и есть тетраподизация (хотя к выходу на сушу она в данном случае не привела). Кроме ризодонтид по пути тетраподизации пошли еще две группы кистеперых. Одна из них — семейство Tristichopteridae, к которому относится позднедевонский эустеноптерон, самая изученная кистеперая рыба на свете, не исключая, пожалуй, и современной латимерии. Но эустеноптерон гораздо менее «тетраподизирован», чем относящаяся к той же эволюционной линии удивительная рыба мандагерия (Mandageria). Другая ветвь, выбравшая схожее направление эволюции, — это тетраподы и близкие к ним амфибиотические рыбы, такие как пандерихт и тиктаалик. Последний был очень похож на настоящую амфибию, но тетраподных лап у него нет. А как, собственно, появились тетраподные лапы? В нормальном рыбьем плавнике есть две части: мясистая основа, имеющая внутренний костный скелет, и перепончатая каемка, скелет которой образован кожными лучами — лепидотрихиями. У некоторых рыб мясистая часть плавников в ходе эволюции постепенно редуцировалась, и осталась только перепончатая каемка с кожными лучами. Так обстоит дело почти у всех лучеперых, за это и получивших свое название, которые составляют подавляющее большинство современных рыб. У тетрапод произошло ровно обратное. Михаил Феодосьевич Ивахненко, очень талантливый палеонтолог, в основном занимавшийся немного другими темами, высказал однажды идею, что ключевым моментом в становлении тетраподной конечности была редукция кожных лучей, за которой последовало компенсаторное разрастание мясистой части. Если Ивахненко прав, то переходной формой между плавником и лапой был ласт. Отличие ласта от плавника в том, что он полностью мускулистый, без перепончатой каемки. Полное исчезновение лепидотрихий мгновенно превращает плавник в ласт, управляемая гибкость которого может пригодиться, если нужно ползать по дну или пробираться сквозь заросли. Эволюционная биология развития подтвердила предположение Ивахненко. Оказалось, что существуют регуляторные гены, которые необходимы для развития кожного скелета парных плавников и которые есть у всех рыб, но отсутствуют у тетрапод. Потеря этих генов, видимо, и была тем самым ключевым событием.
 Еще одно изображение ризодуса. Здесь лучше, чем на главной картинке, видно, что парные плавники у него имеют довольно широкую, укрепленную кожными лучами перепончатую каемку. Рисунок с сайта deviantart.com У ризодонтид ничего подобного не происходит. Их плавники остаются плавниками. Лепидотрихии не только не исчезают — они разрастаются, укрепляются и дополнительно интегрируются с костным скелетом плавника, чтобы создать единую прочную конструкцию. Это принципиально иной путь эволюции конечностей по сравнению с тем, что выбрали тетраподы. Путь, оказавшийся тупиковым: на сушу ризодонтиды не вышли и в конце концов вообще вымерли. Тетраподы, впрочем, тоже вышли на сушу далеко не сразу. Девонские тетраподы (ихтиостега, акантостега, тулерпетон) были существами либо водными, либо в лучшем случае амфибиотическими, но точно не наземными. Первые истинно наземные тетраподы, опорно-двигательная система которых позволяла им не только плавать по мелководью, помогая себе лапами, но и регулярно ходить по суше, появились только в карбоне. Карбон стал эпохой расцвета амфибий — лягушачьим раем, по выражению чудесной писательницы Ирины Яковлевой. Тут произошла парадоксальная вещь. Амфибии, которые вернулись в воду (либо никогда оттуда и не уходили), оказались успешными конкурентами кистеперых рыб в их родной адаптивной зоне. Они неплохо плавали, обладали более эффективным, чем у рыб, легочным дыханием и были в высшей степени универсальными хищниками, способными атаковать любую добычу — от мелкой, вроде членистоногих, до такой, которая сравнима с самим хищником по размеру. Кистеперые рыбы по большинству параметров проигрывали своим потомкам. «Эволюция не может быть справедливой», как однажды сказал Фридрих фон Хайек. Между тем эволюцию земной биосферы никто не останавливал. В карбоне появились деревья с мощной корневой системой, которые «начали подниматься на плакор по долинам рек, удерживая от эрозии берега». Реки и озера стали более стабильными, более глубокими, более прозрачными, с более высоким содержанием кислорода, уже не требующим для поддержания жизни органов воздушного дыхания. Свидетельством этих глобальных перемен служит появление в континентальных водоемах двустворчатых моллюсков. Как известно, эти малоподвижные существа питаются за счет фильтрации воды и дышат только жабрами. Ни падения концентрации кислорода, ни тем более пересыхания водоема они не переносят. Первые пресноводные двустворки появились в конце среднего девона, одновременно с первыми на Земле лесами. До этого их не было. Зато в карбоне их стало еще больше. От перемен больше всех выиграли лучеперые рыбы, которые возникли одновременно с мясистолопастными, но первое время были не слишком успешны. В новых условиях они отказались от легких в пользу плавательного пузыря, отказались от мясистых плавников в пользу чисто перепончатых (которые мы и видим у большинства современных рыб), и всё это дало им решающее преимущество — непревзойденную маневренность. Кистеперые по сравнению с ними были неповоротливы. Кроме того, отказ от легких позволил лучеперым рыбам упразднить смешивание артериальной и венозной крови, которое было неизбежно при сочетании легочного дыхания с жаберным, но порождало чрезвычайно серьезные физиологические ограничения. Конкурировать с безлегочными лучеперыми кистеперые опять же не могли. Зона их обитания неуклонно сужалась, теснимая с двух сторон. Вот тут-то и пробил час ризодонтид. Первые находки ризодонтид относятся к позднему девону, но расцвета они достигают именно в карбоне. Их чудовищное увеличение размера было отчаянной попыткой вырваться из той экологической «узости», в которую эволюция загнала пресноводных кистеперых. Нельзя не признать, что эта попытка в какой-то мере удалась. Правда, успех был ограниченным. Экологическая ниша, которую заняли карбоновые ризодонтиды, больше всего напоминала экологическую нишу современных крупных крокодилов — с тем отличием, что крокодил при желании способен ходить и даже бегать по суше, а у ризодонтид такой возможности не было. Они были универсальными суперхищниками, хватавшими всё живое как в воде, так и на кромке берега. Весьма вероятно, что их излюбленными объектами были другие хищники, стоящие ниже в пищевой цепочке. Для крупных ризодонтид никакая добыча не была слишком велика. К тому же они, подобно акулам и тем же крокодилам, умели отрывать куски от добычи, которую нельзя было сразу проглотить (об адаптации к таким нагрузкам свидетельствует строение их плечевого пояса). Несомненно, среди жертв ризодонтид были не только рыбы, но и амфибии, которым они, можно сказать, от лица всех кистеперых рыб отплатили.
 Это не ризодус, а другая рыба из семейства ризодонтид — австралийская Barameda, которая жила чуть раньше (но тоже в карбоне) и достигала шестиметрового размера. Пишут, что укорененные «клыки» взрослой барамеды имеют 22 сантиметра в длину. На картинке барамеда атакует какую-то крупную амфибию: сцена, которая вполне могла разыграться в карбоновой реке или озере. Рисунок с сайта en.wikipedia.org Закончилась эта «битва богов и титанов» печально для последних. Тут возникает еще один парадокс: выстоять под давлением амфибий ризодонтиды смогли только благодаря конвергенции с ними. На эту конвергенцию обратила внимание Махала Эндрюс (Mahala Andrews), автор, пожалуй, самого полного исследования биологии ризодонтид. В сущности, это тот же процесс, который мы называем тетраподизацией. Но с точки зрения настоящих тетрапод комплекс признаков, который сумели выработать ризодонтиды, был несовершенен. Например, дыхательная система у них, насколько можно судить, осталась полностью рыбьей. А никаких уникальных преимуществ (кроме размера) они не приобрели. Слишком крупные, со слабым хвостом, ризодонтиды навсегда упустили шанс стать хорошими пловцами. Архаичная и в то же время переусложненная конструкция парных плавников не допускала возможности превратить их в наземные лапы. Между тем разнообразие амфибий продолжало неумолимо расти. В начале перми оно достигло пика, и тогда же все оставшиеся пресноводные кистеперые вымерли. Разумеется, история на этом не завершилась. Прошло время, и амфибии, в свою очередь, удалились в тень под натиском новых хозяев суши — рептилий. Ризодонтиды успели застать их появление: первые рептилии (например, Hylonomus) были как раз современниками рода Rhizodus. Но до очередной революции в сообществах, которая произошла в конце перми, наши герои уже не дожили. 12-11-2021 | Просмотров: 823
Комментарии
Комментировать
Комментировать
|

Ещё по теме
|
|
|